Тақырыбы: Холестериннің алмасуы.Майлар алмасуының бұзылуы мен реттелуі.
Тақырыбы: Холестериннің алмасуы. Майлар алмасуының бұзылуы мен реттелуі.
Сабақтың мақсаты:
- Метаболизм және холестериннің алмасуының реттелу механизмі туралы түсінікті қалыптастыру;
2.Кетонды денелердің алмасуы туралы түсінікті қалыптастыру;
3.Қандағы майлардың тасымалдаушы түрі туралы түсікті қалыптастыру.
Оқыту мақсаты:
Биохимиялық құрылым туралы, холестерин алмасу және кетонды денелердің қанның липопротеиндерінің құрылымының ерекшеліктері негізінде май алмасудың бұзылуына байланысты пайда болатын: атеросклероз, гиперлипопротеинемия,кетономия және т.б. аурулардың биохимиялық механизмдерін түсіндіруді үйрену.
Тақырыптың негізгі сұрақтары:
- Холестерин: құрылымы, физиологиялық рөлі, қайнар көздері, организмдегі қолдану жолдары.
- Организмдегі холестериннің биосинтезінің кезеңдері. Осы процестердің реттелуі.
- Холестериннің тасымалдануы және липопротеиндердің осы процестегі рөлі.
- Липопротеиндердің класстары. Атерогенді және антиатерогенді липопротеиндер.
- Холестериннің алмасуының реттелу механизмі. Атеросклероз. Гиперхолестеринемия. Өттегі тас ауруы. Семіздік.
- Кетонды денелердің синтезі және олардың қолданылуы. Кетонемиялар, асқынуының себептері.
- Майлардың алмасуындағы горманальды реттелуі.
- Майлар мен көмірсулардың алмасуының өзара байланысы.
Оқытудың және сабақ берудің әдісі: практикалық оқыту – аз топтардағы жұмыс
Тапсырмаларды келесі түрде бөлу:
1 топ — Холестерин синтезі
2 топ — Кетонды денелер синтезі
3 топ — Холестерин алмасудың реттелу механизмі
4 топ — Липопротеиндердің кластары, функциялары
5 топ –Майлар алмасуынын гормональді реттелуі, бұзылыстары.
Жұмыс барысында келесіге назар аудару:
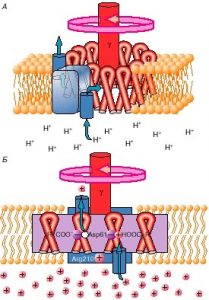
Қан плазмасының барлық майлардың қан тасымалының қамтамасыз ететін суда ерігіш жиынтығы болып табылады, әр класты липопротеидтердің құрамына кіретініне назар аудару керек.
Липопротеиндер тығыздығы үлкейген сайын диаметрі азаятын сфералық бөлшектер болып табылады. Бөлшектердің ядросы полярлы емес липидтері — триацилглицерины, бос және эстерифицирлі холестерин, ал қан плазмасымен байланыста тұратын сыртқы бөлігі ақуызбен (апопротеином) байланысты афолепидті 2 қабат болып табылады. Липопотеидтер және физика- химиялық қасиеті бойынша өте гетерогенді. Липопротеиндер липидтерінің тасымалдаушы түрлері болып табылатын 5 клиникалық ең маңызды процестері бар екеніне назар аудару керек:
Хиломикроны (ХМ) экзогенді триацилглицерин және холестериннің сорылу процесінде ішектің қабырғасында пайда болады. Алдымен ХМ кеудедегі лимфатикалык тармаққа, одан кейін қан тармағына түседі.ХМ экзогенді триацилглицериннің негізгі тасымалдаушы түрі болып табылады.Олардың ХМ кұрамындағы үлесі 81-87%. ХМ электрофорезде катодқа да, анодқа да орын ауыстырмайды, бастауда кала береді.
- Тығыздықтағы өте төмен липопротеиндер (ТӨТЛП) немесе b-липопротеиндер бауырда түзіледі және триацилглицериндердің негізгі тасымалдаушысыболып табылады. Липопротеинограммада хиломикрондармен -b-липопротеинамдер (денедегі электрофорез) арасында орналасқан.
- Төмен тығыздықтагы липопротеиндер (ТТЛП) және b-липопротеидер холестериннің жоғары мөлшерімен (47% дейін) ерекшеленеді және осы липоидтың бауырдан ұлпага негізгі тасымалдау түрі болып табылады. ТТЛП атерогенді ЛП болып табылады.
- Тығыздықтағы жоғары Липопротеиндер (ТЖЛП) a-липопротеиндер бауырда тұз лицитин-холестерин-ацилтрансфераза асерімен құрамындағы холестериннің эстерификация жүзеге асады. Осылайша стерифициленген холестериннің басты тасымалдаушы турі болып табылады. Мұндагы егер ТЖЛП ұлпалар мембраналарына тасымалдаса, онда кері процесс холестерин ұлпа мембранасының жойылуы, ТЖЛП ең негізгі функциясы болып табылып, олардың антирогенді құрылымын камтамасыз етеді.
- Каннын липопротеині канның 99 % ақуыз және 1% эстерифицирленбеген май қышқылдарынан тұратын альбумин май қышқылдары комплексін жатқызуға болады.
Кейбір ауруларда кан сарысуындағы липопротейдтерінің спектрі өзгеруі мүмкін.
Көміркышқылдың шектеулі пайдаланылуын және триацилглицериндерді күшейтілген мобилизациясы кезінде метаболизмнің ацетил — КоА – лимонды кышкылының циклы май қышқылының синтезінің 3 жолының 2 жолының жылдамдығыны айрықша төмендейді. Нәтижесінде ацетил-КоА 3-окси-3метилглутарил-КоА арқылы холестеринге және ацетоацетатқа айналады.
Холестерин – сүтқоректілердің негізгі стерині,манызды биологиялық мәні бар: биологиялык мембрана құрамына кіреді,өт қышқылының, кейбір дәрумендер мен гормондардың негізін қалаушы, сондай-ақ липопротейидтің қажетті кұрамдас болігі болып табылады. Ақуыз холестериннің басты қайнар көзі болып және холестерин плазмасын бөлу орталығы жарты болігі өтке барады, болып кызмет етеді. Бауырдағы холестерин зат алмасу бұзылуы холестерин тастарының, (холестерин кристалдарының) өттің түсуіне келип согады. Қандағы холестерин көбейуі (гиперхолестеринемия) диабет,атеросклероз,менингит және баска ауруларда байқалады, сондықтан қандағы холестеринді анықтау маңызды диагностикалық тест ретінде пайдаланылады. Жалпы холестерин сау адамның плазмасындағы кұрамы 115-247 мг% аралығында тербеледі, оның 60-70% холестерин эфирі болса, ал 30-40 % бос холестерин.
Холестерин организмдегі плазмалық және жасушаішілік мембрананың міндетті құрам бірлігі болып табылады. Оның көп бөлігі миелинді қабатында болады, эритроцит мембранасында және плазмалық мембранада(25% дейін).
Қан құрамындағы холестериннің диагностикалық белгісін анықтауда, мынаған көңіл бөлген жөн, жасқа байланысты холестерин көлемі өспейді. (30-50 жаста холестерин көлемі 150-250мг% құрайды). Тұқымқуалаушылық гиперхолестеринемияда қандаңы холестериннің көлемі 500-1000мгдл жетеді. Қандағы холестериннің төмендеуі (гипохолестеремия) өкпе туберкулезінде, панкретитте, созылмалы жүрек жетіспеушілігінде болады.
Тәжірибелік жұмыста клиникалық- диагностиклық лабароторияда холестеринді анықтау үшін калориметриялық әдіс қолданылады.
Аштықта, ұзақ физикалық күш түскенде және қант диабетінде май қышқылдарының жоғарғы ерігіштігі ацетил-КоА жиналуына әкеледі, ол бүйректе кетон денелеріне айналады. Кетон денелеріне β- гидроксибутират, ацетоацетат және ацетон жатады.
Глюкоза жетіспеушілігінде жасушадағы ҮКҚЦ реакциясының жылдамдығы бауырда төмендейді, өйткені оксалоацетат глюконеогенез үшін қолданылады. Нәтижесінде ацетил-КоА түзілуі жоғарылаған кезде ҮКҚЦ тотығу жылдамдығы мүмкіндігін арттырады. Ацетил-КоА бауыр митохондриясында қорда жинақталады,ол кетон денелерінің синтезінде қолданылады. Кетондық денелердің синтезі тек бауырдың митохондрияларында өтеді, ол ацетил-КоА 2молекуласының байланысуы арқылы басталады, тиолаза әсерімен ацетил-КоА түзіледі. Ацетоацетил-КоА мен ацетил-КоА 3ші молекуласы байланысады да 3-гидрокси-3-метилглутарил-КоА (ГМГ-КоА) түзіледі. Осы реакцияны ГМГ-КоА-синтаза ферменті катализдейді. Ары қарай ГМГ-КоА-лиаза ГМГ-КоАны бос ацетоацетат және ацетил-КоА айналдырады.
Ацетоацетат қанға бөлінуі мүмкін немесе қалпына келу жолымен бауырда β-гидроксибутиратқа айналуы мумкін, өйткені бауыр ұлпаларындағы активті NADH жоғары концентрациясы құрылады. Сондықтан қандағы негізгі кетонды дене β-гидроксибутират.
Ацетоацетаттың жоғары концентрациясы кезінде оның бөлігі ацетонға айналып, ферменті декарбоксиленеді, ацетон ұлпаларымен кайта пайдаланылмайды, бірақ шығарылатын ауамен және зәрмен болініп шығады. Осылайша организм тотығып игермейтін бірак суда еригіш қышқыл болып табылып, ацитоз акелетін кетон денелер шектен тыс колемінен құтылады.
Ұзақ ашыққанда кетонды денелер қанқаның бұлшық етіне жүрек және бауырға энергия көзі болып табылады. Осылайша глюкоза тотығу үшин ми жіне эритроциттерде сакталады. Ашығып бастағаннан кейін 2-3 күннен кейін кет денелердің концентрациясы ми ұлпаларына кіріп оның глюкоза сұранысын азайта отырып тотығуына жеткиликті етеді.β—Гидроксибутират, ұлпаларға жетіп НAD-тәуелді дегидрогеназамен тотыгып және ацетоацетатқа айналады. Ацетоацетат сукцинил-КоА – КоА донорымен күшейеді:
Ацетоацетат + Сукцинил-КоА – Ацетолацетил-КоА + Сукцинат.
Реакцияны сукцинил-КоА-ацетоацетат-КоА-трансфераза катализдейді. Бұл фермент бауырда синтезделмейді, сондықтан бауырдағы кетон денелерді энергия көзі ретінде пайдаланбайды « экспорттайды».
Бақылау:
Тапсырмаларды шешу:
- Есептеу, 3 ацетосірке қышқылының толық тотығуы кезінде неше АТФ молі түзіледі.
- Құрамында олеинхолестерині бар тағаммен жануарларды қоректендіргенде радиоактивтенген бүкіл көмірсу атомы, 2 сағаттан кейін қанның сарысуында хеломикронның табылғанын көреміз. Бірақ бұл тек холестеринде емес оның эфирінде, ТАГ фракциясында да кездеседі. Тәжірибелердің нәтижелерін түсіндіріндер, еске түсіріп және дәптерге жазып:
- a) Тамақтың холестерин майлары жіңішке ішектерде айналады;
- b) Гидролиз кезінде сіңіріп алу процесіне қатысатын компонент, мүше;
- c) Холестерин және май қышқылы эпительді асқазан жасушасында айналады;
- d) Экзогенді холестерин және оның эфиры қанға түскенде липопротеиндері.
- Аэробты тотығу кезіндегі АТФтың 1молекуласының Миристил S СоА дан СО2 ға дейін анықтаныз. Сонымен қатар АТФ молекуласының санын анықтаңыз.
- Толық катаболизмнің схемасын салыңдар олеинды қышқыл, линоленды қышқыл және арахинды қышқыл ацетил S- КоА дейін.
- АТФ тың 6 молекуласы май қышқылының көмірсуынан неге көп.