

1)) Белки и их классификация. Элементарный состав белков. Физико-химические свойства белков. Биологические функции белков. Структурная организация белков. Сложные белки.
Белки – это высокомолекулярные азотсодержащие органические вещества, молекулы которых построены из остатков аминокислот.Св-ва и структура белков: молекулы белков отличаются неисчерпаемым разнообразием структуры при строгой ее специфичности у данного белка; белкам присуща способность к внутримолекулярным взаимодействиям, что обеспечивает динамичность структуры их молекул, изменчивость и пластичность их формы, обратимость переходов из глобулярного состояния в фибриллярное; обладая разнообразными химическими радикалами аминокислотных остатков в составе полипептидных цепей, белковые молекулы способны вступать в разнообразные химические и физические взаимодействия как с друг другом, так и с нуклеиновыми кислотами, полисахаридами, образуя надмолекулярные комплексы; молекулы белков закономерно изменяют свою структуру под влиянием внешнего воздействия и восстанавливают исходное состояние при его снятии многие белки способны каталитически ускорять химические реакции, протекающие в живом организме; белкам присущи регуляторные, защитные, токсические, транспортные, сократительные, структурные, рецепторные и многие другие функцииБелки обладают особой чувствительностью к химическим реагентам (кислоты, щелочи) и легко разрушаются. Белки очень легко теряют свои природные, нативные свойства и переходят в денатурированное состояние. Аминокислоты, находящиеся в белках, связаны друг с другом пептидными связями. Линейная последовательность аминокислот в белке уникальна для каждого индивидуального белка; информация о ней содержится в участке молекулы ДНК, называемой геном. Белки-ферменты(каталитическая функция); сократительные(мышечные) белки(актин,миозин); ядерные белки(гистоновые и негистоновые-регулируют работу генов); белки-гормоны(инсулин-снижает ур-нь глюкозы); защитная ф-я белков(интерферон,фибрин,фибриноген-свертывание крови); структурные белки(коллаген-в соединительных тканях; кератин-в волосах,ногтях;эластин-в кровеносных сосудах); транспортная ф-я(гемоглобин).Белки входят во все основные структурные компоненты клеток, тканей и органов, выполняют ферментативные функции, участвуют в переносе веществ через мембраны. В основе классификации лежат разные принципы:
1)по степени сложности (простые и сложные);
2) по форме молекул (глобулярные и фибриллярные белки);
3)по растворимости в отдельных растворителях (водорастворимые, растворимые в слабых солевых растворах — альбумины, спирторастворимые — проламины, растворимые в щелочах — глютелины),;
4)по выполняемым ими функциям, например запасные белки, скелетные, белки- ферменты, гормоны и т. д.
По степени сложности белки делят на протеины (простые белки), состоящие только из остатков аминокислот, и протеиды (сложные белки), состоящие из белковой (апобелок) и небелковой частей (простетическая группа). Протеины — запасные, скелетные, отдельные ферментные белки. К ним относят:
-альбумины — белки с относительно небольшой молекулярной массой, хорошо растворимые в воде и в слабых солевых растворах; типичный представитель альбуминов- белок яйца — овальбумин;
-глобулины — растворяются в водных растворах солей. Это очень распространенные белки, входят в состав мышечных волокон, крови, молока, они составляют большую часть семян бобовых и масличных культур. Представителем глобулинов животного происхождения является лактоглобулин молока; к глобулинам принадлежат глобулины крови и мышечный белок миозин.
-проламины — растворяются в 60—80 %-ном растворе этилового, спирта. Это характерные белки семян злаков, например: глиадин — пшеницы и ржи, зеин — кукурузы, авенин — овса, гордеин — ячменя;
-глютелины — растворяются только в растворах щелочей. Из них следует выделить оризенин из семян риса и глютенин клейковинных белков пшеницы.
Протеиды — из этой группы сложных белков отметим только следующие:
-нуклеопротеиды — кроме белка включают нуклеиновые кислоты. Нуклеиновые кислоты относятся к важнейшим биополимерам, которым принадлежит огромная роль в наследственности;
-липопротеиды — содержат кроме белка липиды. Содержатся в протоплазме и мембранах. Принимают участие в формировании клейковинных белков;
-фосфопротеиды — кроме белка присутствует фосфорная кислота. Им принадлежит важная роль в питании молодого организма. Пример: казеин — белок молока.
Белки выполняют множество самых разнообразных функций, характерных для живых организмов: каталитическая(ферменты), регуляторная(гормоны), транспортная(гемоглобин), защитная(свертывание крови, иммуноглобулины), сократительная(актин и миозин), структурная(коллаген, кератин).
Структурная организация белков ( в лекции)
Протеины — запасные, скелетные, отдельные ферментные белки. К ним относят:
-альбумины — белки с относительно небольшой молекулярной массой, хорошо растворимые в воде и в слабых солевых растворах; типичный представитель альбуминов- белок яйца — овальбумин;
-глобулины — растворяются в водных растворах солей. Это очень распространенные белки, входят в состав мышечных волокон, крови, молока, они составляют большую часть семян бобовых и масличных культур. Представителем глобулинов животного происхождения является лактоглобулин молока; к глобулинам принадлежат глобулины крови и мышечный белок миозин.
-проламины — растворяются в 60—80 %-ном растворе этилового, спирта. Это характерные белки семян злаков, например: глиадин — пшеницы и ржи, зеин — кукурузы, авенин — овса, гордеин — ячменя;
-глютелины — растворяются только в растворах щелочей. Из них следует выделить оризенин из семян риса и глютенин клейковинных белков пшеницы.
Протеиды — из этой группы сложных белков отметим только следующие:
-нуклеопротеиды — кроме белка включают нуклеиновые кислоты. Нуклеиновые кислоты относятся к важнейшим биополимерам, которым принадлежит огромная роль в наследственности;
-липопротеиды — содержат кроме белка липиды. Содержатся в протоплазме и мембранах. Принимают участие в формировании клейковинных белков;
-фосфопротеиды — кроме белка присутствует фосфорная кислота. Им принадлежит важная роль в питании молодого организма. Пример: казеин — белок молока.
2)) Ферменты, их классификация и номенклатура. Особенности ферментативного катализа, влияние температуры, pH и концентрации субстрата. Регуляция активности ферментов. Изоферменты. Диагностическое значение определения ферментов. Ферментопатии.
По этому принципу все ферменты делят на 6 классов.
- Оксидоредуктазы — ускоряют реакции окисления — восстановления.
- Трансферазы — ускоряют реакции переноса функциональных групп и молекулярных остатков.
- Гидролазы — ускоряют реакции гидролитического распада.
- Лиазы — ускоряют негидролитическое отщепление от субстратов определенных групп атомов с образованием двойной связи (или присоединяют группы атомов по двойной связи).
- Изомеразы — ускоряют пространственные или структурные перестройки в пределах одной молекулы. 6. Лигазы — ускоряют реакции синтеза, сопряженные с распадом богатых энергией связей.
ИЗОФЕРМЕНТЫ (изоэнзимы, изозимы)- это различные по аминокислотной последовательности изоформы или изотипы одного и того же фермента, существующие в одном организме, но, как правило, в разных его клетках, тканях или органах; -это ферменты, катализирующие идентичные реакции, но отличающиеся друг от друга строением и каталитических свойствами. К изоферментам относят только те формы ферментов, появление которых связано с генетически детерминированными различиями в первичной структуре пептидной цепи. Все изоферменты одного и того же фермента выполняют одну и ту же каталитическую функцию, но могут значительно различаться по степени каталитической активности, по особенностям регуляции или другим свойствам. Примером фермента, имеющего изоферменты, является гексокиназа, имеющая четыре изотипа, обозначаемых римскими цифрами от I до IV. При этом один из изотипов гексокиназы, а именно гексокиназа IV, экспрессируется почти исключительно в печени и обладает особыми физиологическими свойствами, в частности её активность не угнетается продуктом её реакции глюкозо-6-фосфатом. Ещё одним примером фермента, имеющего изоферменты, является амилаза — панкреатическая амилаза отличается по аминокислотной последовательности и свойствам от амилазы слюнных желёз, кишечника и других органов. Это послужило основой для разработки и применения более надёжного метода диагностики острого панкреатита путём определения не общей амилазы плазмы крови, а именно панкреатической изоамилазы.
(Остальное в лекции)
3)) Особенности структуры ДНК и РНК, типы РНК. Биосинтез ДНК и РНК. Понятие об опероне. Ингибиторы транскрипции.
Биосинтез ДНК(репликация).Биологический генетический код.
Синтез ДНК протекает в ядре в S-фазу клеточного цикла и предшествует делению клеток. Первоначально клетка из состояния покоя Go вступает в G1-фазу, в ходе которой синтезируются ферменты и белки, необходимые для синтеза ДНК. Затем в S-фазу протекает репликация и диплоидная клетка превращается в тетраплоидную, а в ходе митоза делится, образуя 2 дочерние диплоидные клетки. В эукариотических клетках репликация начинается одновременно во многих участках ДНК, которые имеют специфическую нуклеотидную последовательность и называются ориджинами репликации. От каждого ориджина синтез новых цепей ДНК идет в двух противоположных направлениях, образуя две репликативные вилки. Процесс является полуконсервативным, так как по завершении репликации каждая дочерняя молекула ДНК содержит одну родительскую нить и одну вновь синтезированную. Матрицей служат обе нити ДНК. Репликация включает стадии инициации, элонгации и терминации. В ходе инициации образуются две репликативные вилки при участии ферментов ДНК-топоизомеразы, ДНК-хеликазы и белков, связывающихся с одноцепочечными участками ДНК (SSВ-белки). ДНК-топоизомераза 1 присоединяется к участку ориджина, расщепляет одну из цепей ДНК и связывается с фосфатным остатком в точке разрыва, происходит локальное раскручивание двухцепочечной нити ДНК. Две молекулы ДНК-хеликазы, используя энергию АТФ, разрывают водородные связи между комплементарными основаниями и разделяют цепи ДНК. Одновременно ДНК-топоизомераза восстанавливает фосфодиэфирную связь и освобождается из связи с ДНК. SSВ-белки присоединяются к одноцепочечным участкам и препятствуют их повторному скручиванию в двойную спираль. На стадии элонгации образуются дочерние цепи ДНК на материнской ДНК. Этот процесс катализирует ДНК-полимераза.Сначала ДНК-полимераза синтезирует РНК –праймер, которым начинается лидирующая цепь и каждый фрагмент Оказаки в отстающей нити ДНК. Лидирующая нить растет непрерывно, а отстающая – в виде фрагментов Оказаки, каждый их которых включает включает РНК-праймер (10 нуклеотидов) и участок ДНК, примерно равный длине ДНК в составе нуклеосомы (примерно 150 нуклеотидов). Когда следующий фрагмент Оказаки достигает праймера предыдущего фрагмента, ДНК-полимераза отделяется от синтезированной цепи, а праймер предыдущего фрагмента удаляют эндонуклеаза и РНКаза, образуется брешь. ДНК-полимераза удлиняет последний фрагмент Оказаки, заполняя брешь. ДНК-лигаза сшивает предыдущий и вновь синтезированный фрагменты между собой. Новые цепи синтезируются неодинаково. Одна цепь на матрице ДНК с направлением от 3’- к 5’- концу растет непрерывно по ходу движения репликативной вилки и называется лидирующей.Вторая на матрице с направлением от 5’- к 3’- концу синтезируется против движения репликативной вилки в виде коротких фрагментов – фрагментов Оказаки, ее называют запаздывающей или отстающей. ДНК-лигаза объединяет фрагменты в полинуклеотидную цепь, затрачивая молекулу АТФ на образование каждой 3’, 5’- фосфодиэфирной связи. Кофактором всех стадий репликации являются ионы Mg2+. В результате образуются дочерние цепи, комплементарные и антипараллельные нитям материнской ДНК. После деления каждая дочерняя клетка получает диплоидный набор хромосом, идентичный материнской клетке.Завершение синтеза ДНК в процессе репликации происходит на стадии терминации. Существует система репарации- система восстановления поврежденной молекулы ДНК,которая включает: специфическая эндонуклеаза(обнаруживает нарушение комплементарности и гидролизует 3’,5’-фосфодиэфирную связь в поврежденной нити ДНК); экзонуклеаза(удаляет от 20 до 30 нуклеотидных остатков в области разрыва к 3’- концу образовавшейся бреши и заполняет брешь); ДНК – лигаза(используя АТФ как источник энергии, соединяет 3’,5’-фосфодиэфирной связью место разрыва между вновь синтезированной и основной нитями ДНК).
Генети ческий код — свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов.
Свойства генетического кода:
-триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон).
-непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно.
-неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки).
-однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA у Euplotes crassus кодирует две аминокислоты — цистеин и селеноцистеин)[1]
-вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов.
-универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусов до человека (на этом основаны методы генной инженерии; есть ряд исключений, показанный в таблице раздела «Вариации стандартного генетического кода» ниже).
Биосинтез РНК(транскрипция). РНК-полимеразы. Типы РНК, их роль.
Транскрипция – это синтез РНК на матрице ДНК. Процесс катализируют РНК-полимеразы, которые подобно ДНК-полимеразам, образуют фосфодиэфирные связи между рибонуклеотидами в соответствии с принципами комплементарности к одной из нитей ДНК, которую обозначают как матричную. У эукариот синтез РНК происходит в ядре и митохондриях практически постоянно вне зависимости от фаз клеточного цикла. В ядре РНК синтезируют 3 фермента: РНК-полимераза I катализирует образование рРНК, РНК-полимераза II –синтез мРНК, РНК-полимераза III – образование тРНК. Нуклеотидтрифосфаты (АТФ, ГТФ, ЦТФ, УТФ) выполняют функции субстратов синтеза и источников энергии. В ходе транскрипции матрицей является нить ДНК, имеющая направление от 3’ к 5’-концу, так как все РНК-полимеразы осуществляют рост новых цепей РНК в направлении от 5’ к 3’-концу антипараллельно матрице. Процесс транскрипции включает стадии инициации, элонгации и терминации. РНК-полимеразы узнают место начала транскрипции – промотер, имеющий специфическую последовательность нуклеотидов –ТАТА-. На стадии инициации к –ТАТА-последовательности матричной цепи ДНК присоединяется белок –ТАТА-фактор, который стимулирует присоединение к ДНК РНК- полимеразы и белковых факторов инициации транскрипции. Образующийся комплекс вызывает расплетение двойной нити ДНК длиной в один виток спирали (около 10 нуклеотидных пар).На этапе элонгации происходит удаление факторов инициации и присоединение фактора элонгации. Синтез РНК осуществляется на матричной нити ДНК по принципу комплементарновти. При этом в активном центре РНК-полимеразы каждый последующий нуклеотид связывается с 3’-концом предыдущего нуклеотида. По мере движения РНК-полимеразы по нити ДНК к освободившемуся промотору присоединяются новые молекулы фермента, поэтому один ген может одновременно транскрибироваться несколькими молекулами РНК-полимеразы.Стадия терминации начинается, когда РНК-полимераза достигает специфической последовательности нуклеотидов – сайта терминации. При этом фактор элонгации отделяется от РНК-полимеразы, а фактор терминации присоединяется. Он облегчает отделение синтезированной молекулы пре-РНК и фермента от матрицы ДНК. Молекулы РНК, которые синтезируются РНК-полимеразами, функционально неактивны и являются молекулами-предшественниками – пре-РНК. Они превращаются в зрелые молекулы только после соответствующих посттранскрипционных модификаций – созревания молекул РНК. Установлено, что эукариотические ДНК состоят из участков, кодирующих последовательность аминокислот в отдельных доменах молекулы белка – экзонов и участков, не содержащих информацию о строении белка – интронов. В ходе транскрипции получаются пре-РНК, содержащие участки, комплеменарные экзонам и интронам. В процессе созревания мРНК интроны удаляются, а экзоны соединяются между собой с высокой точностью при помощи комплексов из малых ядерных рибонуклеопротеинов (мяРНП) – сплайсосом. Этот процесс получил название сплайсинга.В клетках синтезируется около 20 семейств тРНК.Представители каждого семейства способны связываться только с одной из 20 аминокислот, входящих в состав белков.
Виды РНК:
– матричная (информационная) РНК(служит посредником при передаче информации, закодированной в ДНК к рибосомам);
– рибосо мные РНК(рРНК) –( несколько молекул РНК, составляющих основу рибосомы. Основной функцией рРНК является осуществление процесса трансляции — считывания информации с мРНК при помощи адапторных молекул тРНК и катализ образования пептидных связей между присоединёнными к тРНК аминокислотами);
– транспортная РНК, тРНК(функцией является транспортировка аминокислот к месту синтеза белка);
4)) Биосинтез белков, этапы трансляции. Регуляция биосинтеза белков. Индукция и экспрессия генов. Антибиотики – ингибиторы матричных биосинтезов.
Процесс биосинтеза белка включает два этапа: транскрипцию и трансляцию.Трансляция (от лат. translatio — перевод) — синтез полипептидной цепи с использованием мРНК в роли матрицы.В трансляции участвуют все три типа РНК:
мРНК служит информационной матрицей;
тРНК доставляют аминокислоты и узнают кодоны;
рРНК вместе с белками образуют рибосомы, которые удерживают мРНК;
тРНК и белок и осуществляют синтез полипептидной цепи.
мРНК транслируется не одной, а одновременно несколькими (до 80) рибосомами. Такие группы рибосом называются полирибосомами (полисомами). На включение одной аминокислоты в полипептидную цепь необходима энергия четырех АТФ.
После окончания трансляции процесс образования белков обычно еще не завершен. Для того чтобы белок приобрел присущие ему функциональные свойства, полипептидная цепь должна определенным образом свернуться в пространстве, сформировав функционально активную («нативную») структуру. Процесс формирования пространственной структуры белка называется фолдингом.Стадии фолдинга:Пространственная (третичная) структура белка характеризуется сочетанием элементов вторичной структуры (α-спиралей, β-слоев), а также гибких участков полипептидной цепи, называемых петлями. Процесс сворачивания полипептидной цепи имеет ступенчатый характер (рис. 43) и включает:1) формирование элементов вторичной структуры; 2) специфическую ассоциацию некоторых элементов вторичной структуры с образованием супервторичной структуры: это могут быть сочетания нескольких α-спиралей, β-слоев или смешанные комплексы данных элементов;3) промежуточную стадию, связанную с формированием основных элементов третичной структуры и образованием гидрофобного ядра молекулы. Молекула приобретает пространственную структуру, близкую к структуре нативного белка. Однако она еще не обладает присущей данному белку функциональной активностью. Это состояние, получившее название «расплавленная глобула», отличается от нативного меньшей степенью упорядоченности структуры (рис. 44);4) образование нативной третичной структуры белка.Факторы фолдинга опеспечивают оптимальные условия для реализации быстрого и эффективного образования нативной пространственной структуры. Выделяют две группы факторов:1. Фолдазы – ферменты фолдинга (белки с каталитической активностью). Как и все ферменты, фолдазы необходимы лишь в каталитических количествах, т. е. в концентрациях, на порядки меньших, чем у «обслуживаемых» ими белков.
- Молекулярные шапероны. Требуются в количествах, близких к стехиометрическим, т. е. сравнимых по величине с концентрацией сворачиваемых белков. Как и фолдазы, не входят в состав конечных продуктов фолдинга.Часто построенная на рибосоме полипептидная цепь не может принять окончательную биологически активную (нативную) конформацию, пока не подвергнется процессингу, или ковалентной модификации. При этом может происходить отщепление ферментами инициирующих аминокислотных остатков, введение в определенные аминокислотные остатки фосфатных, метильных, карбоксильных, олигосахаридных или простетических групп. Например, гормоны поджелудочной железы инсулин и глюкагон синтезируются в виде неактивных предшественников. Сначала из них удаляются сигнальные пептиды, в результате чего образуются прогормоны. Затем уже в секреторных гранулах прогормоны превращаются в активные гормоны.
5)) Введение в обмен веществ. Понятие о метаболизме и его этапах. Анаболические и катаболические пути. Общие принципы регуляции метаболизма. Конечные продукты метаболизма, пути их выведения.
Метаболи зм (от греч. μεταβολή — «превращение, изменение»), или обмен веществ — набор химических реакций, которые возникают в живом организме для поддержания жизни. Эти процессы позволяют организмам расти и размножаться, сохранять свои структуры и отвечать на воздействия окружающей среды. Метаболизм обычно делят на две стадии: в ходе катаболизма сложные органические вещества деградируют до более простых; в процессах анаболизма с затратами энергии синтезируются такие вещества, как белки, сахара, липиды и нуклеиновые кислоты. Органические вещества представлены в основном аминокислотами, углеводами, липидами и нуклеиновыми кислотами. Катаболизмом называют метаболические процессы, при которых расщепляются относительно крупные органические молекулы сахаров, жиров, аминокислот. В ходе катаболизма образуются более простые органические молекулы, необходимые для реакций анаболизма (биосинтеза). Анаболизм — совокупность метаболических процессов биосинтеза сложных молекул с затратой энергии. Сложные молекулы, входящие в состав клеточных структур, синтезируются последовательно из более простых предшественников. В метаболическом пути происходит саморегуляция на уровне субстрата или продукта; например, уменьшение количества продукта может компенсировано увеличить поток субстрата реакции по данному пути.
6)) Строение и биологические функции биомембран. Состав и строение липидного бислоя. Мембранные белки, их функции. Лизосомальные болезни.
Основу мембраны составляет липидный бислой – двойной слой молекул липидов, которые обладают свойствомамфифильности (содержат как гидрофильные, так и гидрофобные функциональные группы). В липидном бислое гидрофобные участки молекул взаимодействуют между собой, а гидрофильные участки обращены в окружающую водную среду.По выполняемым функциям белки в составе мембран делятся на
структурные;
каталитические;
рецепторные;
транспортные.
7)) Биоэнергетика. Аутотрофы и гетеротрофы. Основные энергетические субстраты клеток. Эндэргонические и экзэргонические реакции в живых системах. Макроэргические соединения.
Биоэнергетика — раздел биохимии, изучающий энергетические процессы в клетке.Автотрофы – это организмы, способные строить свои тела за счет неорганических соединений, используя солнечную энергию.К ним относятся растения ( только растения). Они синтезируют из СО, НО (неорганические молекулы) под воздействием солнечной энергии – глюкозу (органические молекулы) и О. Они составляют первое звено в пищевой цепи и находятся на 1 трофическом уровне.
Гетеротрофы – это организмы, которые не могут строить собственное тело из неорганических соединений, а вынуждены использовать созданное автотрофами, употребляя их в пищу.К ним относятся консументы и детритофаги. И находятся на II и выше трофическом уровне. Человек тоже гетеротроф.
Вернадскому принадлежит идея, что возможно превращение человеческого общества из гетеротрофного и автотрофное.
В силу своих биологических особенностей человек не может перейти к автотрофности, но общество в целом способно осуществить автотрофный способ производства пищи, т.е. замена природных соединений (белки, жиры, углеводы) на органические соединения, синтезированные из неорганических молекул или атомов.основные органические вещества, поступающие с пищей и входящие в состав организма человека, окисляются с образованием АТФ и потому могут служить источниками энергии. Главные из них — белки, липиды и углеводы. Эндергонические реакции – это химические реакции, требующие притока энергии для их осуществления. В этих реакциях изменение свободной энергии AG положительная величина.Экзергонические реакции – это реакции, в которых энергия выделяется, ᴛ.ᴇ. они идут с выделением энергии. В таких реакциях изменение свободной энергии AG-отрицательная величина.
Внутриклеточные химические реакции бывают представлены в виде˸
- катаболических (экзергонических) реакций;
- анаболических (эндергонических) реакций.
- Понятие о тканевом дыхании по биологическому окислению. Дегидрирование субстратов и окисление водорода как источник энергии в клетке. Локализация митохондриальной цепи переноса электронов.
Тканевое дыхание и биологическое окисление. Распад органических соединений в живых тканях, сопровождающийся потреблением молекулярного кислорода и приводящий к выделению углекислого газа и воды и образованию биологических видов энергии, называется тканевым дыханием. Тканевое дыхание представляют как конечный этап пути превращений моносахаров (в основном глюкозы) до указанных конечных продуктов, в который на разных стадиях включаются другие сахара и их производные, а также промежуточные продукты распада липидов(жирные кислоты), белков (аминокислоты) и нуклеиновых оснований. Итоговая реакция тканевого дыхания будет выглядеть следующим образом:
С6Н12О6 + 6O2 = 6СO2+ 6Н2O + 2780 кДж/моль. (1)
Потребление кислорода тканями зависит от интенсивности реакций тканевого дыхания. Наибольшей скоростью тканевого дыхания характеризуются почки, мозг, печень, наименьшей – кожа, мышечная ткань (в покое). Уравнение (2) описывает суммарный результат многоступенчатого процесса, приводящего к образованию молочной кислоты(см. главу 10) и протекающего без участия кислорода:
С6Н12Об = 2С3Н6О3 + 65 кДж/моль. (2)
ТФ – чрезвычайно важная молекула клетки. Она непрерывно синтезируется и используется. Энергия для синтеза АТФ поступает от субстратов, содержащих высокоэнергетические электроны в ходе их дегидрирования. Электроны высоких энергий извлекаются из субстратов в ходе гликолиза (в цитозоле), при преобразовании пирувата в ацетил-КоА и в цикле трикарбоновых кислот (в митохондриаль-ном матриксе). Молекулы НАДН и ФАДН2 переносят эти электроны в дыхательную цепь, локализованную в во внутренней митохондриальной мембране.
Дыхательная цепь состоит из переносчиков электронов, сгруппированных в 4 ферментативных комплекса, и ферментов. Особенность переносчиков электронов дыхательной цепи в том, что каждый способен принимать электроны от предыдущего и отдавать последующему. Прием и передача электронов молекулами переносчиков происходит упорядоченно в соответствии с их электрохимическими потенциалами. Под электрохимическим потенциалом молекулы понимают её способность вступать во взаимодействие с электроном. Т.е. присоединять электрон и при этом восстанавливаться и отдавать его и при этом окисляться. Электрохимический потенциал молекулы это её способность участвовать в окислительно-восстановительных реакциях. В начало дыхательной цепи электроны вносит НАДН – он обладает самым низким электрохимическим потенциалам. В конце дыхательной цепи находиться кислород – его электрохимический потенциал самый высокий. Электроны, перемещаясь по дыхательной цепи в сторону кислорода, совершают полезную работу по переносу протонов из митохондриалыуого матрикса в межмембранное пространство (рис. 13). На восходящих участках тра-ектории каждый электрон перемещается вместе с протоном, т.е. в форме атома водорода, затем отделяется от протона на внешней стороне мембраны и продолжает движение внутри мембраны. В межмембранном пространстве накапливаются протоны из митохондриального матрикса. Электрон теряет порции своей энергии на всех участках траектории. На восходящих участках траектории электрон расходует энергию на совершение полезной работы, а на нисходящих участках полезная работа не совершается, хотя электрон также теряет часть своей энергии. 40-45 % энергии электронов используется для создания протонного градиента, который, в свою очередь расходуется на синтез АТФ. 20-25 % энергии электронов используется для транспорта необходимых субстратов. Остальная энергия электронов рассеивается в виде тепла, поэтому митохондрии место теплообразования. Движение электронов по дыхательной цепи создает протонный градиент и значительную трансмембранный электрохимический потенциал между межмембранным пространством и митохондриальным матриксом (его обозначают АцН+ = 0,25 Вольта). ДцН* складывается из ДЧ* = 0,2 Вольта (это электрический потенциал, обусловленный разными зарядами по обе стороны мембраны) и АН+ = 0,05 Вольта (химический потенциал, обусловленный различной концентрацией протонов по обе стороны мембраны).
- Механизмы трансформации энергии в клетке (теория П. Митчелла). Окислительное фосфорилирование, коэффициенты Р:О и АДФ:АТФ.
учение о механизме преобразования энергии в биол. мембранах при синтезе аденозинтрифосфорной к-ты (АТФ). Разработана П. Митчеллом в 1961—66. Согласно исходным представлениям Митчелла, запасание энергии в АТФ происходит вследствие предварит, накопления зарядов на стенках мембраны, создания мембранного потенциала и разности концентраций протонов. Разность электрохимич. потенциалов ионов водорода на сопрягающих мембранах (внутр. мембраны митохондрий, тилакоиды хлоропластов, мембраны бактерий) возникает за счёт энергии, выделяемой при деятельности цепи окислит.-восстановит, ферментов, или за счёт поглощённых квантов света. Трансмембранные электрохимич. потенциалы ионов могут служить источником энергии не только для синтеза АТФ, на и для транспорта веществ, движения бактериальных клеток и др. энергозависимых процессов.
Согласно предложенной П. Митчелом гипотезе, движущей силой фосфорилирования АДФ служит энергия разности редокс-потенциалов, возникающая при переносе электронов от окисляемого субстрата по дыхательной цепи к кислороду и трансформируемая в энергию протонного электрохимического потенцила ( треугольник и мю H+).
1.Ферменты дых. Цепи расположены в строго определенной последовательности : каждый последующий белок обладает большим сродством к электронам, чем предыдущий (он более электроположителен, т.е. обладает более положительным окислительно-восстановительным потенциалом). Это обеспечивает однонаправленное движение электронов.
- Все атомы водорода, отщепленные дегидрогеназами от субстратов в аэробных условиях, достигают внутренней мембраны митохондрий в составе НАДН или ФАДН2.
3.Здесь атомы водорода (от НАДН и ФАДН2) передают свои электроны в дыхательную цепь , по которой электроны движутся (50-200 шт/сек )к своему конечному акцептору- кислороду. В результате образуется вода.
4.Поступающие в дыхательную цепь электроны богаты свободной энергией. По мере их продвижения по цепи они теряют энергию.
Часть энергии электронов используется в 1,3,4 комплексами дыхательных ферментов для перемещения ионов водорода через мембрану в межмембранное пространство. Другая часть рассеивается в виде тепла. Упрощенно сказанное представить в виде равенства:
Изменение энергии электронов=совершение работы+ выделение тепла
- Перенос ионов водорода через мембрану (выкачивание) происходит неслучайно , а в строго определенных участках мембраны. Эти участки называются участки сопряжения (или, не совсем точно, пункты фосорилирования). Они представлены 1,3,4 комплексами дыхательных ферментов. В результате работы этих комплексов формируется градиент ионов водорода между внутренней и наружной поверхностями внутренней митохондриальной мембраны. Такой градиент обладает потенциальной энергией.
- 1 Понятие об общих путях катаболизма
Если процесс катаболизма рассматривать с общей точки зрения, то можно выделить три основные его части: Расщепление в пищеварительном тракте. Это гидролитические реакции, превращающие сложные пищевые вещества в относительно небольшое число простых метаболитов: глюкоза, аминокислоты, глицерин, жирные кислоты.
- Специфические пути катаболизма. Простые метаболиты подвергаются специфическим реакциям расщепления, в результате которых образуется либо пировиноградная кислота, либо ацетил – СоА. Причем ацетил – СоА может образоваться из пирувата в результате окислительного декарбоксилирования. Могут также образоваться другие соединения, непосредственно включающиеся в цитратный цикл.
- Цитратный цикл и дыхательная цепьзавершают расщепление пищевых веществ до конечных продуктов – СО2и Н2О.
Следовательно, начиная со стадии образования пирувата происходит унификация путей катаболизма. Из большого числа исходных соединений образуется всего два – пируват и ацетил – СоА. Процесс, начинающийся от пирувата, называется общим путем катаболизма и в свою очередь включает:
- окислительное декарбоксилирование пирувата
- цитратный цикл.
Именно в общем пути катаболизма образуется основная масса субстратов для реакций дегидрирования. Совместно с дыхательной цепью и окислительным фосфорилированием общий путь катаболизма является основным источником энергии в форме АТР.
Окислительное декарбоксилирование пировиноградной кислоты
Суммарный результат многостадийной реакции выглядит следующим образом:
Реакция катализируется тремя ферментами, работающими в определенной последовательности и объединенными в пируватдегидрогеназный комплекс:
Этот комплекс ферментов работает подобно конвейеру, в котором продукт передается от фермента к ферменту. Такой принцип повышает эффективность работы ферментов, так как снижает случайность в контакте реагирующих веществ с ферментом. Далее приводятся названия ферментов и характеристика катализируемых реакций.
- Пируватдекарбоксилаза (1). В качестве кофермента в реакции участвует тиаминдифосфат – производное витамина В1. Фермент катализирует отщепление карбоксильной группы в виде СО2, а ацетильный остаток присоединяет к липоевой кислоте – коферменту второго фермента. Получается ацетил-липоат.
- Дигидролипоат-ацетилтрансфераза(2) – второй фермент комплекса. Катализирует перенос ацетильного остатка, соединенного с липоевой кислотой на второй кофермент HS-СоА с образованием ацетил-СоА. Таким образом, в этой реакции участвуют два кофермента: липоевая кислота, прочно соединенная с ферментом, и кофермент А, объединяющийся с ферментом в момент реакции. Водород остается связанным с липоевой кислотой, которая превращается в дигидролипоат.
- Дегидрогеназа дигидролипоевой кислоты (3) отщепляет водород от липоевой кислоты и переносит его на NAD+. Далее водород транспортируется дыхательной цепью
Главные продукты реакции – это NADH+H+ и ацетил-СоА. NADH+H+ далее окисляется в дыхательной цепи, где энергия используется на синтез 3 моль АТР, а ацетил-СоА окисляется в цитратном цикле. Пируватдекарбоксилазный комплекс находится на внутренней мембране митохондрий и соединен с ней со стороны матрикса.
Цитратный цикл
Цитратный цикл (цикл Кребса, цикл трикарбоновых кислот) – это система реакций, приводящая к полному окислению двухуглеродного ацетильного фрагмента, имеющего различное происхождение. Цитратный цикл является общим конечным путем окисления белков, жиров и углеводов. Все реакции цитратного цикла, как и окислительного декарбоксилирования пирувата, локализованы в митохондриях. В ходе одного полного цикла происходит:
- полное окисление ацетильного остатка до двух молекул СО2;
- образование трех молекул восстановленного NAD+и одной молекулы FADH2;
- образование одной молекулы GTP в результате субстратного фосфорилирования.
Реакции цитратного цикла, ферменты и их характеристика приведена на рисунке:
Схема цитратного цикла; ферменты: 1- пируватдегидрогеназный комплекс, 2- цитратсинтаза, 3- аконитаза, 4- изоцитратдегидрогеназа, 5- a –кетоглутаратдегидрогеназный комплекс, 6- сукцинил-КоА-тиокиназа, 7- сукцинатдегидрогеназа, 8- фумараза, 9- малатдегидрогеназа
Сопряжение общих путей катаболизма с дыхательной цепью
В общих путях катаболизма происходит пять реакций дегидрирования: одна на стадии окислительного декарбоксилирования пирувата и четыре в цитратном цикле. Все 10 атомов водорода переносятся на коферменты дегидрогеназ, которые в свою очередь окисляются в дыхательной цепи. Окисленные коферменты возвращаются в реакции общих путей катаболизма. Регенерация коферментов – это обязательное условие для протекания реакции дегидрирования. Таким образом, общий путь катаболизма и дыхательная цепь непрерывно связаны между собой и отдельно функционировать не могут.
Энергетика цитратного цикла и общих путей катаболизма
За один оборот цитратного цикла синтезируется 12 молекул АТР. Девять из них образуются за счет энергии транспорта в дыхательной цепи трех пар водорода от трех молекул NADH + H+. Две молекулы АТР синтезируются при окислении 1 молекулы FADH2, так как в дыхательной цепи в данном случае действуют только два пункта сопряжения с окислительным фосфорилированием ADP. Кроме того, в цитратном цикле происходит одна реакция субстратного фосфорилирования, дающая 1 моль GTP (АТР). В общих путях катаболизма синтезируется 15 молекул АТР. Три из них при окислительном декарбоксилировании пирувата и 12 – в цитратном цикле.
Регуляция общих путей катаболизма
Главным фактором, регулирующим скорость дыхания и фосфорилирования, являются энергетические потребности организма. Основная масса восстановленных эквивалентов для дыхательной цепи поступает из общих путей катаболизма. Следовательно, регуляция общих путей катаболизма и дыхательной цепи тесно связана. Все контролирующие механизмы осуществляются на уровне ферментов и многие из них с помощью аллостерических эффекторов. Для оценки энергетического состояния клетки используют величину энергетического заряда, отражающего соотношение концентрации ATP к продуктам ее распада – ADP и AMP. При увеличении энергетического заряда в клетке (в состоянии покоя) скорость реакций общих путей катаболизма снижается, а при уменьшении энергетического заряда – увеличивается. Это достигается тем, что ATP действует как аллостерический ингибитор, а ADP и AMP – как аллостерические активаторы некоторых ферментов:
Реакции цитратного цикла и регуляция общего пути катаболизма
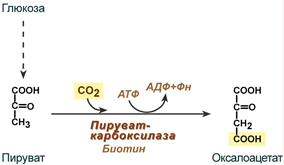
Другой механизм регуляции связан с необходимостью регенерации NAD+ в дыхательной цепи. При уменьшении расхода АТР в клетке скорость дыхания митохондрий снижается (дыхательный контроль), уменьшается также скорость окисления NADH в дыхательной цепи и увеличивается концентрация NADH. В этом случае NADH ингибирует некоторые ферменты общих путей катаболизма, что приводит к замедлению реакций катаболизма и, следовательно, замедлению наработки восстановленных коферментов и уменьшению синтеза АТР. При увеличении энергетических потребностей организма происходит все наоборот. Ряд промежуточных продуктов цитратного цикла служат предшественниками для синтеза необходимых организму веществ. Так сукцинил-СоА используется для синтеза гема, оксалоацетат и a -кетоглутарат – для синтеза аспарагиновой и глутаминовой кислот. Очевидно, что выведение хотя бы одного метаболита нарушает работу цикла, так как уменьшает регенерацию оксалоацетата. Для компенсации концентрации метаболитов цикла в митохондриях происходит реакция карбоксилирования пирувата с образованием оксалоацетата. Таким образом, пируват включается в цитратный цикл двумя путями: окислительное декарбоксилирование с образованием ацетил-СоА, карбоксилирование с образованием оксалоацетата. Последнюю реакцию катализирует пируваткарбоксилаза, коферментом является биотин:
Гипоэнергетические состояния
Наиболее частой причиной гипоэнергетических состояний является гипоксия, возникновение которой в свою очередь связано с нарушением:
- поступления кислорода в кровь, что наблюдается при недостаточности О2во вдыхаемом воздухе или нарушении легочной вентиляции;
- транспорта кислорода в тканипри нарушении кровообращения или снижении транспортной функции гемоглобина;
- функций митоходрий, вызванное действием ядов, разобщителей.
Кроме того, причиной гипоэнергетических состояний могут быть гиповитаминозы, так как в реакциях общих путей катаболизма и дыхательной цепи участвуют коферменты, содержащие витамины. Так, витамин В1 входит в состав тиаминдифосфата, В2 является составной частью FMN и FAD, витамин РР в виде никотинамида входит в состав NAD+ и NADP+, пантотеновая кислота – в состав кофермента А, биотин также выполняет коферментную функцию активации СО2.
11.Цикл лимонной кислоты.. Связь общих путей катаболизма с цепью переноса электронов и протонов. Аллостерические механизмы регуляции цитратного цикла.
Цикл трикарбоновых кислот
Цикл протекает в матриксе митохондрий и представляет собой восемь последовательных реакций.
В первой реакции связываются ацетил и оксалоацетат (щавелевоуксусная кислота) с образованием цитрата (лимонной кислоты), далее происходит изомеризация лимонной кислоты до изоцитрата и две реакции окисления с сопутствующим выделением СО2 и восстановлением НАД.
В пятой реакции образуется ГТФ, это реакция субстратного фосфорилирования.
Последние три реакции составляют так называемый мотив, т.е. набор однотипных преобразований некой молекулы. В данном случае мотив представляет собой окисление, гидратацию и новое окисление, и используется для введения кетогруппы в структуру сукцината.
В итоге после восьми реакций цикла вновь образуется оксалоацетат.
Указанный мотив (ФАД-зависимое окисление, гидратация и НАД-зависимое окисление) также присутствует в реакциях β-окисления жирных кислот. В обратной последовательности (восстановление, дегидратация и восстановление) этот мотив наблюдается в реакциях синтеза жирных кислот.
Основная роль ЦТК заключается в
- генерации атомов водородадля работы дыхательной цепи, а именно трех молекул НАДН и одной молекулы ФАДН2.
Кроме этого, в ЦТК образуется
- одна молекулаГТФ, которая равнозначнаАТФ,
- сукцинил-SКоА, участвующий в синтезе гема,
- кетокислоты, являющиеся аналогами аминокислот – α-кетоглутарат для глутаминовой кислоты, оксалоацетат для аспарагиновой.
Регуляция цикла трикарбоновых кислот
Главным и основным регулятором ЦТК является оксалоацетат, а точнее его доступность. Наличие оксалоацетата вовлекает в ЦТК ацетил-SКоА и запускает процесс.
Обычно в клетке имеется баланс между образованием ацетил-SКоА (из глюкозы, жирных кислот или аминокислот) и количеством оксалоацетата. Источником оксалоацетата являетсяглюкоза (синтез из пирувата в анаплеротической реакции), поступление из фруктовых кислотсамого цикла (яблочной, лимонной), образование из аспарагиновой кислоты в результатетрансаминирования.
{kind=link}
Примером существенной роли оксалоацетата служит активация синтеза кетоновых тел икетоацидоз плазмы крови при недостаточном количестве оксалоацетата в печени. Такое состояние наблюдается при инсулинзависимом сахарном диабете, при голодании, алкогольном отравлении или длительной физической нагрузке.
- Катаболизм глюкозы. Анаэробный и аэробный распад глюкозы.
Гликолиз – это серия реакций, в результате которых глюкоза распадается на две молекулы пирувата (аэробный гликолиз) или две молекулы лактата (анаэробный гликолиз). Все десять реакций гликолиза протекают в цитозоле и характерны для всех органов и тканей. Аэробный распад глюкозы включает реакции аэробного гликолиза и последующее окисление пирувата в реакциях катаболизма.
Таким образом, аэробный распад глюкозы – это предельное ее окисление до СО2 и Н2О, а анаэробный гликолиз – это специфический путь катаболизма, то есть часть аэробного распада глюкозы. Анаэробный распад включает те же реакции специфического пути распада глюкозы до пирувата, но с последующим превращением пирувата в лактат (то есть термины анаэробный распад и анаэробный гликолиз совпадают). Последовательность реакций гликолиза приведена на рисунке:
В гликолизе можно выделить три основных этапа. На первом этапе превращениям подвергаются гексозы, на втором – триозы, на третьем – карбоновые кислоты. Характеристика гликолиза:
- большинство реакций обратимо, за исключением трех (реакций 1, 3, 10);
- все метаболиты находятся в фосфорилированной форме;
- источником фосфатной группы в реакциях фосфорилирования являются АТР (реакции 1, 3) или неорганический фосфат (реакция 6);
- регенерация NAD+, являющаяся необходимым условием протекания гликолиза, происходит при аэробном гликолизе посредством дыхательной цепи. В этом случае водород транспортируется в митохондрии с помощью челночного механизма при участии переносчиков. Это происходит потому, что мембрана митоходрий непроницаема для протонов. При анаэробном гликолизе регенерации NAD+осуществляется независимо от дыхательной цепи. В этом случае акцептором водорода от NADH является пируват, который восстанавливается в лактат;образование АТР при гликолизе может идти двумя путями: либо субстратным фосфорилированием, когда для фосфорилирования ADP используется энергия макроэргической связи субстрата (реакции 7, 9), либо путем окислительного фосфорилирования ADP, сопряженного с дыхательной цепью (реакция 6).В гликолизе можно выделить три основных этапа. На первом этапе превращениям подвергаются гексозы, на втором – триозы, на третьем – карбоновые кислоты. Характеристика гликолиза:
В аэробном гликолизе образуется 10 моль АТР на 1 моль глюкозы. Так, в реакциях 7, 10 образуется 4 моль АТР путем субстратного фосфорилирования, а в реакции 6 синтезируется 6 моль АТР (на 2 моль глицероальдегидфосфата) путем окислительного фосфорилирования:
Суммарный эффект аэробного гликолиза составляет 8 моль АТР, так как в реакциях 1 и 3 используется 2 моль АТР. Дальнейшее окисление двух моль пируват в общих путях катаболизма сопровождается синтезом 30 моль АТР ( по 15 моль на каждую молекулу пирувата . Следовательно, суммарный энергетический эффект аэробного распада глюкозы до конечных продуктов составляет 38 моль АТР.
{kind=link}
Значение анаэробного гликолиза
Анаэробный и аэробный гликолиз энергетически неравноценны. Образование двух моль лактата из глюкозы сопровождается синтезом всего двух моль АТР , потому что NADH, полученный при окислении глицероальдегидфосфата, не используется дыхательной цепью, а акцептируется пируватом.
Анаэробный гликолиз, несмотря на небольшой энергетический эффект, является основным источником энергии для скелетных мышц в начальном периоде интенсивной работы, то есть в условиях, когда снабжение кислородом ограничено. Кроме того, зрелые эритроциты извлекают энергию за счет анаэробного окисления глюкозы, потому что не имеют митохондрий. 9))
- Биосинтез глюкозы – глюконеогенез. Взаимосвязь гликолиза и глюконеогенеза (цикл Кори).
Некоторые ткани, например мозг, нуждаются в постоянном поступлении глюкозы. Когда поступление углеводов в составе пищи недостаточно, содержание глюкозы в крови некоторое время поддерживается в пределах нормы за счёт расщепления гликогена в печени. Однако запасы гликогена в печени невелики. Они значительно уменьшаются к 6-10 ч голодания и практически полностью исчерпываются после суточного голодания. В этом случае в печени начинается синтез глюкозы de novo – глюконеогенез. Глюконеогенез – процесс синтеза глюкозы из веществ неуглеводной природы. Его основной функцией является поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок. Процесс протекает в основном в печени и менее интенсивно в корковом веществе почек, а также в слизистой оболочке кишечника. Эти ткани могут обеспечивать синтез 80-100 г глюкозы в сутки. На долю мозга при голодании приходится большая часть потребности организма в глюкозе. Это объясняется тем, что клетки мозга не способны, в отличие от других тканей, обеспечивать потребности в энергии за счёт окисления жирных кислот (см. раздел 8).
Кроме мозга, в глюкозе нуждаются ткани и клетки, в которых аэробный путь распада невозможен или ограничен, например эритроциты (они лишены митохондрий), клетки сетчатки, мозгового слоя надпочечников и др.
Первичные субстраты глюконеогенеза – лактат, аминокислоты и глицерол. Включение этих субстратов в глюконеогенез зависит от физиологического состояния организма.
Реакции глюконеогенеза
Большинство реакций глюконеогенеза протекает за счёт обратимых реакций гликолиза (рис. 7-45, реакции 9, 8, 7, 6, 5, 4, 2) и катализируется теми же ферментами. Однако 3 реакции гликолиза термодинамически необратимы. На этих стадиях реакции глюконеогенеза протекают другими путями.
Необходимо отметить, что гликолиз протекает в цитозоле, а часть реакций глюконеогенеза происходит в митохондриях. Образование фосфоенолпирувата из пирувата происходит в ходе двух реакций (рис. 7-45, реакции 11, 12), первая из которых протекает в митохондриях. Пируват, образующийся из лактата или из некоторых аминокислот, транспортируется в матрикс митохондрий и там карбоксилируется с образованием оксалоацетата Пируват-карбоксилаза, катализирующая данную реакцию, – митохондриальный фермент, коферментом которого является биотин. Реакция протекает с использованием АТФ.
Дальнейшие превращения оксалоацетата протекают в цитозоле. Следовательно, на этом этапе должна существовать система транспорта оксалоацетата через митохондриальную мембрану, которая для него непроницаема. Оксалоацетат в митохондриальном матриксе восстанавливается с образованием маната (рис. 7-47) при участии NADH (обратная реакция цитратного цикла). Образовавшийся малат затем проходит через митохондриальную мембрану с помощью специальных переносчиков. Кроме того, оксалоацетат способен транспортироваться из митохондрий в цитозоль в виде аспартата в ходе малат-аспартатного челночного механизма Цикл Кори — совокупность биохимических ферментативныхпроцессов транспорта лактата из мышц в печень, и дальнейшего синтеза глюкозы из лактата,катализируемое ферментами глюконеогенеза.
Биологический смысл
При интенсивной мышечной работе, а также в условиях отсутствия или недостаточного числа митохондрий(например, в эритроцитах или мышцах) глюкоза вступает на путь анаэробного гликолиза с образованиемлактата. Лактат не может далее окисляться, он накапливается (при его накоплении в мышцах раздражаютсячувствительные нервные окончания, что вызывает характерное жжение в мышцах). С током крови лактатпоступает в печень. Печень является основным местом скопления ферментов глюконеогенеза (синтезглюкозы из неуглеводных соеднений), и лактат идет на синтез глюкозы.
Реакция превращения лактата в пируват катализируется лактатдегидрогеназой, далее пируватподвергается окислительному декарбоксилированию или может подвергаться брожению.
- Пентозофосфатный путь глюкозы
Это путь превращения глюкозы в пентозы. В пентозофосфатном пути превращения глюкозы можно выделить две части: А – окислительный путь и Б – неокислительный путь синтеза пентоз. Коферментом дегидрогеназ является NADP+, который восстанавливается в NADPH и используется клетками в реакции восстановления и гидроксилирования. Кроме того, пентозофосфатный путь (окислительный и неокислительный) поставляет клетке пентозофосфаты, необходимые для синтеза нуклеиновых кислот и коферментов (NAD, FAD, СоА).
Все реакции пентозофосфатного пути проходят в цитозоле клетки. Реакции неокислительного этапа пентозофосфатного пути являются обратимыми, поэтому становится возможным синтез гексоз из пентоз. Некоторые метаболиты неокислительного пути являются также и метаболитами гликолиза. Из этого следует, что оба процесса тесно связаны и в зависимости от потребностей клетки возможны переключения с одного пути на другой. При сбалансированной потребности в NADPH и пентозах в клетке происходит окислительный путь синтеза пентоз. Если потребности в пентозах превышают потребности в NADPH, то окислительный путь шунтируется за счет использования метаболитов гликолиза: фруктозо-6-фосфат и глицероальдегидфосфат в реакциях неокислительного пути превращаются в пентозы. Если же NADPH необходим в большей степени, чем пентозы, то возможны два варианта:
- при высоком энергетическом статусе клетки излишки пентоз путем обратных реакций неокислительного пути превращаются в фруктозо-6-фосфат и глицероальдегидфосфат, из которых в процессе глюконеогенеза образуется глюкоза;
- при низком энергетическом статусе клетки из пентоз также образуются глицероальдегидфосфат и фруктозо-6-фосфат, которые затем включаются в гликолиз
Наиболее активно реакции пентозофосфатного пути идут в цитозоле клеток печени, жировой ткани, эритроцитах, коре надпочечников, молочной железе при лактации, в гораздо меньшей степени в скелетных мышцах. Этот путь окисления глюкозы не связан с образованием энергии, а обеспечивает анаболизм клеток. В связи с этим у новорожденных и детей первых лет жизни его активность довольно высока.
Пентозофосфатный путь включает два этапа – окислительный и структурных перестроек
| На первом, окислительном, этапе глюкозо-6-фосфат в трех реакциях превращается в рибулозо-5-фосфат, реакции сопровождаются восстановлением двух молекул НАДФ до НАДФН. На этом этапе происходит регуляция процесса: инсулинповышает активность глюкозо-6-фосфат-дегидрогеназы ифосфоглюконат-дегидрогеназы. |
- Строение и функции гликогена. Биосинтез и распад гликогена. Нарушения обмена гликогена. Гликогенозы и агликогенозы. Роль адреналина и глюкагона в мобилизации гликогена. Аллостерическая регуляция гликогенфосфорилазы.
Гликоген – разветвлённый гомополимер глюкозы, в котором остатки глюкозы соединены в линейных участках α-1,4-гликозидной связью. В точках ветвления мономеры соединены α-1,6-гликозидными связями. Эти связи образуются примерно с каждым десятым остатком глюкозы. Следовательно, точки ветвления в гликогене встречаются примерно через каждые десять остатков глюкозы. Таким образом, в молекуле гликогена имеется только одна свободная аномерная ОН-группа и, следовательно, только один восстанавливающий (редуцирующий) конец.
В клетках животных гликоген – основной резервный полисахарид. При полимеризации глюкозы снижается растворимость образующейся молекулы гликогена и, следовательно, её влияние на осмотическое давление в клетке.
Гликоген хранится в цитозоле клетки в форме гранул диаметром 10-40 нм. С гранулами связаны и некоторые ферменты, участвующие в метаболизме гликогена, что облегчает их взаимодействие с субстратом. Разветвлённая структура гликогена обусловливает большое количество концевых мономеров, что способствует работе ферментов, отщепляющих или присоединяющих мономеры при распаде или синтезе гликогена, так как эти ферменты могут одновременно работать на нескольких ветвях молекулы. Гликоген депонируется главным образом в печени и скелетных мышцах.
После приёма пищи, богатой углеводами, запас гликогена в печени может составлять примерно 5% от её массы. В мышцах запасается около 1% гликогена, однако масса мышечной ткани значительно больше и поэтому общее количество гликогена в мышцах в 2 раза больше, чем в печени. Гликоген может синтезироваться во многих клетках, например в нейронах, макрофагах, клетках жировой ткани, но содержание его в этих тканях незначительно. В организме может содержаться до 450 г гликогена.
Распад гликогена печени служит в основном для поддержания уровня глюкозы в крови в постабсорбтивном периоде. Поэтому содержание гликогена в печени изменяется в зависимости от ритма питания. При длительном голодании оно снижается почти до нуля. Гликоген мышц служит резервом глюкозы – источника энергии при мышечном сокращении. Мышечный гликоген не используется для поддержания уровня глюкозы в крови. Распад гликогена или его мобилизация происходят в ответ на повышение потребности организма в глюкозе. Гликоген печени распадается в основном в интервалах между приёмами пищи, кроме того, этот процесс в печени и мышцах ускоряется во время физической работы.
Распад гликогена происходит путём последовательного отщепления остатков глюкозы в виде глюкозо-1-фосфата.
Расход гликогена в мышцах зависит в основном от физической нагрузки
Синтез гликогена
Гликоген синтезируется в период пищеварения (через 1-2 ч после приёма углеводной пищи). Следует отметить, что синтез гликогена из глюкозы (рис. 7-23), как и любой анаболический процесс, является эндергоническим, т.е. требующим затрат энергии.
Регуляция обмена углеводов осуществляется нейроэндокринным путем. Основная роль принадлежит гипоталамической области, гипофизу (АКТГ, тиреотропный, соматотропный гормоны), бета-клеткам островков поджелудочной железы (инсулин), надпочечникам (глюкокортикоиды, адреналин) и щитовидной железе.
Нарушения содержания гликогена проявляются в уменьшении или увеличении количества его в тканях и появлении там, где он обычно не выявляется. Эти нарушения наиболее ярко выражены при сахарном диабете и при наследственных углеводных дистрофиях – гликогенозах.
При сахарном диабете, развитие которого связывают с патологией бета-клеток островков поджелудочной железы, что обусловливает недостаточную выработку инсулина, происходит недостаточное использование глюкозы тканями, увеличение ее содержания в крови (гипергликемия) и выведение с мочой (глюкозурия). Тканевые запасы гликогена резко уменьшаются. Это в первую очередь касается печени, в которой нарушается синтез гликогена, что ведет к инфильтрации ее жирами – развиваетсяжировая дистрофия печени; при этом в ядрах гепатоцитов появляются включения гликогена, они становятся светлыми
Глюкагон – “гормон голода”, вырабатываемый α-клетками поджелудочной железы в ответ на снижение уровня глюкозы в крови. По химической природе глюкагон – пептид.
Адреналин выделяется из клеток мозгового вещества надпочечников в ответ на сишалы нервной системы, идущие из мозга при возникновении экстремальных ситуаций (например, бегство или борьба), требующих внезапной мышечной деятельности. Адреналин является сигналом “тревоги”. Он должен мгновен В печени существует и аллостерическая регуляция гликогенфосфорилазы: ее ингибирует АТФ и глюкозо-6ф, а активирует АМФ.но обеспечить мышцы и мозг источником энергии.
- Пищевые жиры и их переваривание. Роль желчных кислот. Ресинтез жиров в кишечнике. Хиломикроны. Нарушение переваривания и всасывания липидов.
С пищей в организм ежедневно поступает от 80 до 150 г липидов. Основную массу составляют жиры, наряду с глюкозой служащие главными источниками энергии. Хотя калорийность жиров значительно выше, чем углеводов (9 по сравнению с 4,7 ккал/моль), при рациональном питании жиры обеспечивают не более 30% от общего количества калорий, поступающих с пищей. Жидкие жиры (масла) содержат в своём составе полиеновые жирные кислоты, которые не синтезируются в организме; поэтому жидкие жиры должны составлять не менее одной трети жиров пищи. С липидами в организм поступают и жирорастворимые витамины A, D, Е, К. Переваривание липидов пищи происходит в кишечнике. Основные продукты гидролиза (жирные кислоты и 2-моноацилглицеролы) после всасывания подвергаются ресинтезу и последующей упаковке в хиломикроны (ХМ) в клетках слизистой оболочки кишечника.
Роль желчи
1. Наряду с панкреатическим соком нейтрализация кислого химуса, поступающего из желудка. При этом карбонаты взаимодействуют с НСl, выделяется углекислый газ и происходит разрыхление химуса, что облегчает переваривание.
2. Обеспечивает переваривание жиров,
уменьшает поверхностное натяжение, что препятствует сливанию капель жира,
образование мицелл, способных всасываться.
3. Благодаря п.п.1 и 2 обеспечивает всасывание жирорастворимых витаминов (витамин A, витамин D, витамин K, витамин E). 4. Усиливает перистальтику кишечника.
5. Экскреция избытка ХС, желчных пигментов, креатинина, металлов Zn, Cu, Hg, лекарств. Для холестерина желчь – единственный путь выведения, с ней может выводиться 1-2 г/сут.
Ресинтез липидов – это синтез липидов в стенке кишечника из поступающих сюда экзогенных жиров, иногда могут использоваться и эндогенные жирные кислоты. Основная задача этого процесса – связать поступившие с пищей средне- и длинноцепочечные жирные кислоты со спиртом – глицеролом или холестеролом. Это ликвидирует их детергентное действие на мембраны и позволяет переносить по крови в ткани. Хиломикроны — класс липопротеинов, образующихся в тонком кишечнике в процессе всасывания экзогенных липидов. Это самые большие из липопротеинов, достигающие размера от 75 нм до 1,2 микрона в диаметре. Хиломикроны синтезируются клетками кишечника и секретируются в лимфатические сосуды, после чего попадают в кровь
Переваривание липидов может легко нарушиться
Любое нарушение внешнего обмена липидов (проблемы переваривания или всасывания) проявляется увеличением содержания жира в кале – развивается стеаторея.
Причины нарушений переваривания липидов
1. Снижение желчеобразования в результате недостаточного синтеза желчных кислот и фосфолипидов при болезнях печени, гиповитаминозах.
2. Снижение желчевыделения (обтурационная желтуха, билиарный цирроз, желчнокаменная болезнь). У детей часто причиной может быть перегиб желчного пузыря, который сохраняется и во взрослом состоянии.
3. Снижение переваривания при недостатке панкреатической липазы, который возникает при заболеваниях поджелудочной железы (острый и хронический панкреатит, острый некроз, склероз). Может возникать относительная недостаточность фермента при сниженном выделении желчи.
4. Избыток в пище катионов кальция и магния, которые связывают жирные кислоты, переводят их в нерастворимое состояние и препятствуют их всасыванию. Эти ионы также связывают желчные кислоты, нарушая их работу.
5. Снижение всасывания при повреждении стенки кишечника токсинами, антибиотиками (неомицин, хлортетрациклин).
6. Недостаточность синтеза пищеварительных ферментов и ферментов ресинтеза липидов в энтероцитах при белковой и витаминной недостаточности.
- Обмен жирных кислот. β-окисление высших жирных кислот. Энергетический выход.
Жирные кислоты поступают с пищей или синтезируются в организме Субстраты, необходимые для синтеза жирных кислот, образуются при катаболизме глюкозы и таким образом, часть глюкозы превращается сначала в жирные кислоты, а затем в жиры. Хотя специфический путь катаболизма жирных кислот заканчивается образованием ацетил-КоА, служащим исходным субстратом для синтеза жирных кислот, процессы синтеза и окисления жирных кислот необратимы. Они происходят в разных компартментах клеток (биосинтез протекает в цитозоле, а окисление – в митохондриях) и катализируются разными ферментами. Окисление жирных кислот как источников энергии увеличивается в постабсорбтивный период, при голодании и физической работе. В этих состояниях их концентрация в крови увеличивается в результате мобилизации из жировых депо, и они активно окисляются печенью, мышцами и другими тканями. При голодании часть жирных кислот в печени превращается в другие “топливные” молекулы – кетоновые тела. Они, в отличие от жирных кислот, могут использоваться нервной тканью как источник энергии. При голодании и длительной физической работе кетоновые тела служат источником энергии для мышц и некоторых других тканей.
А. β-Окисление жирных кислот
β-Окисление – специфический путь катаболизма жирных кислот, при котором от карбоксильного конца жирной кислоты последовательно отделяется по 2 атома углерода в виде ацетил-КоА. Метаболический путь – β-окисление – назван так потому, что реакции окисления жирной кислоты. β-Окисление жирных кислот происходит только в аэробных условиях. Процесс собственно β-окисления состоит из 4-х реакций, повторяющихся циклически. В них последовательно происходит окисление (ацил-SКоА-дегидрогеназа), гидратирование(еноил-SКоА-гидратаза) и вновь окисление 3-го атома углерода (гидроксиацил-SКоА-дегидрогеназа). В последней, трансферазной, реакции от жирной кислоты отщепляетсяацетил-SКоА. К оставшейся (укороченной на два углерода) жирной кислоте присоединяется HS-КоА, и она возвращается к первой реакции. Все повторяется до тех пор, пока в последнем цикле не образуются два ацетил-SКоА.
- Биосинтез жирных кислот. Роль цитрата, биотина, ацетил-коА. Структура и биологическая роль пальмитатсинтазы.
С пищей в организм поступают разнообразные жирные кислоты, в том числе и незаменимые. Значительная часть заменимых жирных кислот синтезируется в печени, в меньшей степени – в жировой ткани и лактирующей молочной железе. Источником углерода для синтеза жирных кислот служит ацетил-КоА, образующийся при распаде глюкозы в абсорбтивном периоде. Таким образом, избыток углеводов, поступающих в организм, трансформируется в жирные кислоты, а затем в жиры.
- Синтез пальмитиновой кислоты
Образование ацетил-КоА и его транспорт в цитозоль
Синтез жирных кислот происходит в абсорбтивный период. Активный гликолиз и последующее окислительное декарбоксилирование пирувата способствуют увеличению концентрации ацетил-КоА в матриксе митохондрий. Так как синтез жирных кислот происходит в цитозоле клеток, то ацетил-КоА должен быть транспортирован через внутреннюю мембрану митохондрий в цитозоль. Однако внутренняя мембрана митохондрий непроницаема для ацетил-КоА, поэтому в матриксе митохондрий ацетил-КоА конденсируется с оксалоацетатом с образованием цитрата при участии цитратсинтазы:
Ацетил-КоА + Оксалоацетат -> Цитрат + HS-КоА.
Затем транслоказа переносит цитрат в цитоплазму.
Перенос цитрата в цитоплазму происходит только при увеличении количества цитрата в митохондриях, когда изоцитратдегидрогеназа и α-кетоглутаратдегидрогеназа ингибированы высокими концентрациями NADH и АТФ. Эта ситуация создаётся в абсорбтивном периоде, когда клетка печени получает достаточное количество источников энергии. В цитоплазме цитрат расщепляется под действием фермента цитратлиазы:
Цитрат + HSKoA + АТФ → Ацетил-КоА + АДФ + Pi + Оксалоацетат.
Ацетил-КоА в цитоплазме служит исходным субстратом для синтеза жирных кислот, а окса-лоацетат в цитозоле подвергается следующим превращениям (см. схему ниже).
Пируват транспортируется обратно в матрикс митохондрий. Восстановленный в результате действия малик-фермента NADPH используется как донор водорода для последующих реакций синтеза жирных кислот. Другой источник NADPH – окислительные стадии пентозофосфатного пути катаболизма глюкозы.
Образование малонил-КоА из ацетил-КоА – регуляторная реакция в биосинтезе жирных кислот.
Первая реакция синтеза жирных кислот – превращение ацетил-КоА в малонил-КоА. Фермент, катализирующий эту реакцию (ацетил-КоА-карбоксилаза), относят к классу лигаз. Он содержит ковалентно связанный биотин. Активность фермента ацетил-КоА-карбоксилазы определяет скорость всех последующих реакций синтеза жирных кислот.
Реакции, катализируемые синтазой жирных кислот, – ферментным комплексом, катализирующим реакции синтеза пальмитиновой кислоты,
После образования малонил-КоА синтез жирных кислот продолжается на мультиферментном комплексе – синтазе жирных кислот (пальмитоилсинтетазе). Этот фермент состоит из 2 идентичных протомеров, каждый из которых имеет доменное строение и, соответственно, 7 центров, обладающих разными каталитическими активностями. Этот комплекс последовательно удлиняет радикал жирной кислоты на 2 углеродных атома, донором которых служит ма-лонил-КоА. Конечный продукт работы этого комплекса – пальмитиновая кислота, поэтому прежнее название этого фермента – пальмитоилсинтетаза.
Первая реакция – перенос ацетильной группы ацетил-КоА на тиоловую группу цистеина ацетилтрансацилазным центром. Затем от малонил-КоА остаток малонила переносится на сульфгидрильную группу ацилпереносящего белка малонилтрансацилазным центром. После этого комплекс готов к первому циклу синтеза.
Ацетильная группа конденсируется с остатком малонила по месту отделившегося СО2. Реакция катализируется кетоацилсинтазным центром. Образовавшийся радикал ацетоацетила
последовательно восстанавливается кетоацил-редуктазой, затем дегидратируется и опять восстанавливается еноилредуктазой – активными центрами комплекса. В результате первого цикла реакций образуется радикал бутирила, связанный с субъединицей синтазы жирных кислот.
Суммарное уравнение синтеза пальмитиновой кислоты из ацетил-КоА и малонил-КоА имеет следующий вид:
CH3-CO-SKoA + 7 HOOC-CH2-CO-SKoA + 14 (NADPH + H+) → C15H31COOH + 7 СО2 + 6 Н2О + 8 HSKoA + 14 NADP+.
Рис. 8-38. Синтез пальмитиновой кислоты. Синтаза жирных кислот: в первом протомере SH-группа принадлежит цистеину, во втором – фосфопантетеину. После окончания первого цикла радикал бутирила переносится на SH-группу первого протомера. Затем повторяется та же последовательность реакций, что и в первом цикле. Пальмитоил-Е – остаток пальмитиновой кислоты, связанный с синтазой жирных кислот. В синтезированной жирной кислоте только 2 дистальных атома углерода, обозначенные *, происходят из ацетил-КоА, остальные – из малонил-КоА.
413
Рис. 8-39. Общая схема реакций синтеза пальмитиновой кислоты.
донором водорода в которых служит кофермент NADPH. Восстановление NADP+ происходит в реакциях:
- дегидрирования в окислительных стадиях пентозофосфатного пути катаболизма глюкозы;
- дегидрирования малата малик-ферментом;
- дегидрирования изоцитрата цитозольной NADP-зависимой дегидрогеназой.
- Регуляция синтеза жирных кислот
Регуляторный фермент синтеза жирных кислот – ацетил-КоА-карбоксилаза. Этот фермент регулируется несколькими способами.
- Ассоциация/диссоциация комплексов субъединиц фермента.В неактивной форме ацетил-КоА-карбоксилаза представляет собой отдельные комплексы, каждый из которых состоит из 4 субъединиц. Активатор фермента – цитрат; он стимулирует объединение комплексов, в результате чего активность фермента увеличивается. Ингибитор – пальмитоил-КоА; он вызывает диссоциацию комплекса и снижение активности фермента (рис. 8-40).
- Фосфорилирование/дефосфорилирование ацетил-КоА-карбоксилазы.В постабсорбтивном состоянии или при физической работе глюкагон или адреналин через аденилатциклазную систему активируют протеинкиназу А и стимулируют фосфорилирование субъединиц ацетил-КоА карбоксилазы. Фосфорилированный фермент неактивен, и синтез жирных кислот останавливается. В абсорбтивный период инсулин активирует фосфатазу, и ацетил-КоА карбоксилаза переходит в дефосфорилированное состояние (рис. 8-41). Затем под действием цитрата происходит полимеризация протомеров фермента, и он становится активным. Кроме активации фермента, цитрат выполняет и другую функцию в синтезе жирных кислот. В аб-сорбтивный период в митохондриях клеток печени накапливается цитрат, в составе которого остаток ацетила транспортируется в цитозоль.
- Индукция синтеза ферментов.Длительное потребление богатой углеводами и бедной жирами пищи приводит к увеличению секреции инсулина, который стимулирует индукцию синтеза ферментов: ацетил-КоА-карбоксилазы, синтазы жирных кислот, цитратлиазы,
Рис. 8-41. Регуляция ацетил-КоА-карбоксилазы.
изоцитратдегидрогеназы. Следовательно, избыточное потребление углеводов приводит к ускорению превращения продуктов катаболизма глюкозы в жиры. Голодание или богатая жирами пища приводит к снижению синтеза ферментов и, соответственно, жиров.
19.Биосинтез и мобилизация кетоновых тел. Физиологическая роль. Кетонемия и кетонурия.
При голодании, длительной физической работе и в случаях, когда клетки не получают достаточного количества глюкозы, жирные кислоты используются многими тканями как основной источник энергии. В отличие от других тканей мозг и другие отделы нервной ткани практически не используют жирные кислоты в качестве источника энергии. В печени часть жирных кислот превращается в кетоновые тела, которые окисляются мозгом, нервной тканью, мышцами, обеспечивая достаточное количество энергии для синтеза АТФ и уменьшая потребление глюкозы. К кетоновым телам относят β-гидроксибутират, ацетоацетат и ацетон. Первые две молекулы могут окисляться в тканях, обеспечивая синтез АТФ. Ацетон образуется только при высоких концентрациях кетоновых тел в крови и, выделяясь с мочой, выдыхаемым воздухом и потом, позволяет организму избавляться от избытка кетоновых тел.
Синтез кетоновых тел в печени. При низком соотношении инсулин/глюкагон в крови в жировой ткани активируется распад жиров. Жирные кислоты поступают в печень в большем количестве, чем в норме, поэтому увеличивается скорость β-окисления. Скорость реакций ЦТК в этих условиях снижена, так как оксалоацетат используется для глюконеогенеза. В результате скорость образования ацетил-КоА превышает способность ЦТК окислять его. Ацетил-КоА накапливается в митохондриях печени и используется для синтеза кетоновых тел. Синтез кетоновых тел происходит только в митохондриях печени.
Синтез кетоновых тел начинается с взаимодействия двух молекул ацетил-КоА, которые под действием фермента тиолазы образуют ацетоацетил-КоА (рис. 8-33). С ацетоацетил-КоА взаимодействует третья молекула ацетил-КоА, образуя 3-гидрокси-3-метилглутарил-КоА (ГМГ-КоА). Эту реакцию катализирует фермент ГМГ-КоА-синтаза. Далее ГМГ-КоА-лиаза катализирует расщепление ГМГ-КоА на свободный ацетоацетат и ацетил-КоА.
Ацетоацетат может выделяться в кровь или превращаться в печени в другое кетоновое тело – β-гидроксибутират путём восстановления.
В клетках печени при активном β-окислении создаётся высокая концентрация NADH. Это способствует превращению большей части ацетоацетата в β-гидроксибутират, поэтому основное кетоновое тело в крови – именно β-гидроксибутират. При голодании для многих тканей жирные кислоты и кетоновые тела становятся основными топливными молекулами. Глюкоза используется в первую очередь нервной тканью и эритроцитами.
При высокой концентрации ацетоацетата часть его неферментативно декарбоксилируется, превращаясь в ацетон. Ацетон не утилизируется тканями, но выделяется с выдыхаемым воздухом и мочой. Таким путём организм удаляет избыточное количество кетоновых тел, которые не успевают окисляться, но, являясь водорастворимыми кислотами, вызывают ацидоз.
Рис. 8-33. Синтез кетоновых тел в митохондриях гепатоцитов. Регуляторный фермент синтеза кетоновых тел (ГМГ-КоА-синтаза) ингибируется свободным КоА. – реакция идёт неферментативно при высокой концентрации кетоновых тел в крови.
Регуляция синтеза кетоновых тел. Регуляторный фермент синтеза кетоновых тел – ГМГ-КоА синтаза.
- ГМГ-КоА-синтаза – индуцируемый фермент; его синтез увеличивается при повышении концентрации жирных кислот в крови. Концентрация жирных кислот в крови увеличивается при мобилизации жиров из жировой ткани под действием глюкагона, адреналина, т.е. при голодании или физической работе.
- ГМГ-КоА-синтаза ингибируется высокими концентрациями свободного кофермента А..
Кетоацидоз. В норме концентрация кетоновых тел в крови составляет 1-3 мг/дл (до 0,2 мМ/л), но при голодании значительно увеличивается. Увеличение концентрации кетоновых тел в крови называют кетонемией, выделение кетоновых тел с мочой – кетонурией. Накопление кетоновых тел в организме приводит к кетоацидозу: уменьшению щелочного резерва (компенсированному ацидозу), а в тяжёлых случаях – к сдвигу рН (некомпенсированному ацидозу), так как кетоновые тела (кроме ацетона) являются водорастворимыми органическими кислотами (рК~3,5), способными к диссоциации:
СН3-СО-СН2-СООН ↔ СН3-СО-СН2-СОО– + Н+.
Ацидоз достигает опасных величин при сахарном диабете, так как концентрация кетоновых тел при этом заболевании может доходить до 400-500 мг/дл. Тяжёлая форма ацидоза – одна из основных причин смерти при сахарном диабете. Накопление протонов в крови нарушает связывание кислорода гемоглобином, влияет на ионизацию функциональных групп белков, нарушая их кон-формацию и функцию.
Кетонемия
Кетонемия – повышенное содержание в крови кетоновых тел. Возникает такое состояние при тяжёлой форме сахарного диабета или голодании.
При голодании в крови падает концентрация глюкозы, а при диабете глюкоза не поступает в клетку с необходимой скоростью. В результате начинается усиленный липолиз для высвобождения необходимой энергии. Мобилизованные жировые кислоты направляются из жировых депо в печень, где и образуются кетоновые тела. Пока их количество в пределах нормы, периферические ткани успевают произвести их окисление и получить таким образом недостающую энергию. При превышении нормы скорости окисления не хватает, и кетоны накапливаются в кровотоке.
Кетонемия обычно сопровождается кетонурией – накоплением кетоновых тел в моче. Выявить кетонемию можно по результатам анализовHYPERLINK “http://zoovet.ru/lab.htm” мочи и крови.
20.Обмен стероидов. Биосинтез холестерина. Выведение желчных кислот из организма. Биохимия атеросклероза.
Холестерин является общим предшественником всех стероидных гормонов . В соответствии с этим, обмен холестерина в тканях, секретирующих стероиды, чрезвычайно важен для регуляции биосинтеза стероидных гормонов . Основные процессы обмена холестерина представлены на рис. 1-5сер .
В течение многих лет считалось, что весь или большая часть холестерина, используемого для стероидогенеза синтезируется из ацетата в клетках, продуцирующих стероиды.
Хотя синтез холестерина действительно имеет место в этих клетках, липопротеины плазмы, которые синтезируются в печени , по всей вероятности, являются главным источником холестерина, используемого для стероидогенеза ( рис.1-4сер ).
Относительная важность различных классов липопротеинов как источников холестерина для стероидогенеза зависит от вида животного, но у человека липопротеины низкой плотности (ЛПНП ) играют наиболее важную роль.
Мембранные рецепторы липопротеинов обнаружены в стероид-продуцирующих клетках и связывание липопротеина с рецептором приводит к интернализации комплекса липопротеин-рецептор. Попав в стероидогенную клетку, липопротеины разрушаются лизосомальными ферментами и освобождают холестерин; рецептор может подвергаться далее деградации или возвращается в клеточную мембрану для дальнейшего использования. Захват и деградация липопротеинов регулируется гормонами, так что при увеличении потребности в стероидных гормонах, повышенные количества холестерина как субстрата поступают в стероид-продуцирующие клетки.
Свободный холестерин в стероид-продуцирующих клетках, независимо от того, возник ли он из липопротеинов или синтезировался в клетке, может немедленно использоваться для стероидогенеза или превращаться в эфиры холестерина . Клетки, которые синтезируют стероидные гормоны, содержат большое количество эфиров холестерина в структурах, известных как липидные капли . Эфиры холестерина являются основной формой запасов холестеринов и должны подвергнуться гидролизу до свободного холестерина, который используется в стероидогенезе. Скорость гидролиза эфиров холестерина, также как захват и деградация липопротеинов находится под регулирующим воздействием гормонов. Таким образом, доставка свободного холестерина в стероид-продуцирующие клетки может тесно координироваться с общей активностью процессов стероидогенеза.Холестерин поступает в организм из животной пищи или синтезируется в печени из других компонентов пищи. Подобно другим жирам, холестерин не растворяется в крови (которая имеет водную основу) и для перемещения по кровеносной системе должен прикрепляться к белкам. Существует два типа белков, переносящих холестерин. Белок первого типа, ЛПНП (липопротеин низкой плотности), призван доставлять холестерин к клеткам-потребителям, где он используется по назначению. При контакте ЛПНП с мембраной (оболочкой) клетки, холестерин легко отсоединяется от ЛПНП и проникает в клетку. В клетках существуют рецепторы, которые отвечают за количество поглощенного холестерина.
Соединение ЛПНП-холестерин легко окисляется. Это означает, что он становится легкой добычей свободных радикалов кислорода, и в этот момент сам выступает в роли свободного радикала, способного повреждать стенки кровеносных сосудов. По этой причине очень важно достаточное использование антиоксидантов для профилактики сердечно-сосудистых заболеваний (ССЗ). Кроме этого, имея низкую плотность, ЛПНП легко теряют холестерин, который переходит на стенки сосудов.
ЛПВП (липопротеин высокой плотности), наоборот, имеют свойство “впитывать” свободный холестерин с поверхности клетки, избытки холестерина, а также холестерин, связанный с ЛПНП, и высвобождать его только в печени, выполняя функцию пылесоса, высасывающего холестерин из клеток организма, и затем транспортного средства для доставки его в печень. При этом ЛПВП никогда не теряют холестерин при транспортировке его в печень.
В печени часть холестерина превращается в желчные кислоты, а часть вместе с желчными кислотами поступает в кишечник. В кишечнике холестерин и желчные кислоты частично могут реабсорбироваться (снова всасываться в кишечнике) либо выводиться из организма вместе с калом. Значительную роль в связывании и выведении из организма холестерина и желчных кислот играют некоторые виды растительной клетчатки. Это определяет необходимость достаточного потребления клетчатки для снижения уровня холестерина в крови.
Нормальное содержание холестерина в сыворотке крови взрослого человека – 3,9 – 5,2 ммоль/л.
Более высокий уровень холестерина в крови требует коррекцию диетой, а при еще более значительном повышении необходима медикаментозная коррекция. Большое значение в профилактике атеросклероза имеет содержание ЛПВП в крови. Рекомендуемые цифры для подростков и взрослых – 1,5 – 3,3 г/л.
21.Обмен триацилглицеридов и фосфолипидов в клетках. Ожирение. Липотропные факторы.
ТАГ – депо энергии. Мобилизуются адреналином и глюкагоном при стрессах, голодании, физ.нагр. за счет активации ТАГ-липазы. Жирные кислоты поступают в кровь, связываются с альбумином и поступают в органы. Синтез ТАГ активно протекает в кишечнике, печени, жировой ткани. Необходимы α-глицеролфосфаты (глицерол-3-фосфат) и ацил-КоА.
В-ва, которые усиливают синтез фосфолипидов липотропные. Они способствуют образованию липопротеинов, которые выносят из печени эндогенные жиры, препятствуют жировому перерождению. Липотропные факторы – ненасыщенные ЖК, Витамины В6,Вс,В12; холин. При гидролизе фосфолипидов образуются лизофосфатиды, за счет отщепления ЖК в β-положении. Это мощные детергенты, разрушающие биомембрану и провоцирующие отступление.
Окисление глицерина: глицерин=>глицерол-3-фосфат; глицеролфосфат =>фосфодиоксиацетат. При окислении в аэробных условиях обр-ся 20 или 22 АТФ; в анаэробных условия 1 АТФ и лактат.
Начальные реакции синтеза триацилглицеролов и фосфолипидов совпадают и происходят при наличии глицерола и жирных кислот.
В реакциях биосинтеза можно выделить следующие события:
- Образование глицерол-3-фосфатачерез диоксиацетонфосфат из глюкозы или при фосфорилировании свободного глицерола.
2.. Биосинтез фосфатидной кислоты – требует наличия глицерол-3-фосфата и жирных кислот. При связывании глицерол-3-фосфата с жирными кислотами синтезируется фосфатидная кислота.
Далее фосфатидная кислота может превращаться двумя путями – в ЦДФ-ДАГ или дефосфорилироваться до 1,2-ДАГ.
- Синтез триацилглицерола– идет из 1,2-ДАГ после дефосфорилирования фосфатидной кислоты. Образованный 1,2-ДАГ ацилируется до ТАГ.
- Синтез фосфолипидов. Сейчас рассматриваются два пути синтеза фосфолипидов.
-По одному пути 1,2-ДАГ не превращается в ТАГ, а связывается с этаноламином с образованием фосфатидилэтаноламина, либо с холином – образуется фосфатидилхолин.
-По другому пути, ЦДФ-ДАГ связывается либо с инозитолом, либо с серином с образованием соответственно фосфатидилинозитола или фосфатидилсерина. При декарбоксилировании фосфатидилсерина далее образуется фосфатидилэтаноламин, который может превратиться, в свою очередь, в фосфатидилхолин.
Синтезированный любым способом фосфатидилэтаноламин также способен взаимодействовать с серином и обратно образовывать фосфатидилсерин.
Ожирение – избыточное количество нейтрального жира в подкожной жировой клетчатке. Выделяют два типа ожирения – первичное и вторичное.
Первичное ожирение – Является следствием нарушения баланса энергии в результате гиподинамии и переедания. В здоровом организме количество поглощаемой пищи регулируется гормоном адипоцитов лептином. Лептин вырабатывается в ответ на увеличение жировой массы в клетке и, в конечном итоге, снижая в гипоталамусе образование нейропептида Y (который стимулирует поиск пищи), подавляет пищевое поведение. У 80% лиц с первичным ожирением гипоталамус нечувствителен к лептину, у 20% имеется дефект структуры лептина.
Вторичное ожирение – Возникает при гормональных заболеваниях. К таким заболеваниям относятся, например, гипотиреоз, гиперкортицизм.
Принцип лечения
При лечении ожирения в первую очередь необходимо учитывать тот факт, что любое ожирение – это вопрос баланса, т.е. прихода и расхода энергии. (увеличение физической нагрузки ; диета с резким уменьшением количества сладких продуктов и насыщенных жиров (молочные продукты, сыр, сливочное масло, сало, жирное мясо) ; прием биологически активных пищевых добавок (БАД) подавляющих липогенез, стимулирующих липолиз и окисление жирных кислот (цитримакс, гуарана, L-карнитин), поливитаминных и полиминеральных препаратов при обязательной физической нагрузке; при вторичном ожирении обязательно лечение основного заболевания.)
Липотропные вещества – Все вещества, способствующие синтезу ФЛ и препятствующие синтезу ТАГ, называются липотропными факторами. К ним относятся:
- Структурные компонентыфосфолипидов: полиненасыщенные жирные кислоты, инозитол, серин, холин, этаноламин.
- Метионин– донор метильных групп, полученных в обмене серина и глицина, для синтеза холина и фосфатидилхолина.
- Витамины: пиридоксин(В6), способствующий образованию ФЭА из ФС; цианкобаламин(В12) и фолиевая кислота, участвующие в образовании активной формы метионина, и, следовательно, в синтезе фосфатидилхолина.
22.Переваривание и всасывание белков. Биохимические механизмы регуляции пищеварения. Активация протеаз. Нарушение всасывания аминокислот.
Переваривание белков – многоэтапный процесс, контролируемый пептидазами.
Расщепление белков до аминокислот начинается в желудке, продолжается в двенадцатиперстной кишке и заканчивается в тонком кишечнике. В некоторых случаях распад белков и превращения аминокислот могут происходить также в толстом кишечнике под влиянием микрофлоры.
Протеолитические ферменты подразделяют по особенности их действия на экзопептидазы, отщепляющие концевые аминокислоты, и эндопептидазы, действующие на внутренние пептидные связи.
В желудке начинается переваривание белков. Здесь действуют два основных фермента: ПЕПСИН и ГАСТРИКСИН.
ПЕПСИН вырабатывается главными клетками слизистой желудка в неактивной форме – ПЕПСИНОГЕН, который под действием соляной кислоты активируется и превращается в пепсин. Процесс активации является АУТОКАТАЛИТИЧЕСКИМ путём ЧАСТИЧНОГО ПРОТЕОЛИЗА.
Соляная кислота вырабатывается обкладочными клетками слизистой желудка и играет очень важную роль в переваривании белков:-активирует ПЕПСИНОГЕН, превращая его в пепсин,- создаёт оптимум рН для действия пепсина (1,5 – 2),-обладает бактерицидным действием, -денатурирует белки, которые потом гидролизуются.
Внутренние ПЕПТИДНЫЕ связи становятся доступными для действия пепсина, способствует продвижению желудочного содержимого далее в кишечник.
Пепсин – это фермент, который является ЭНДОПЕПТИДАЗОЙ, т.е. действует на внутренние ПЕПТИДНЫЕ связи, в образовании которых участвуют ароматические аминокислоты (ФЕН, ТИР, ТРИ) своими АМИДНЫМИ группами. ГАСТРИКСИН по действию аналогичен пепсину. – ЭНДОПЕПТИДАЗА. Его оптимум рН = 3 – 3,5. Действует на ПЕПТИДНЫЕ связи. В желудке под действием ПЕПСИНА и ГАСТРИКСИНА сложные белковые молекулы распадаются на высокомолекулярные ПОЛИПЕПТИДЫ. Ими являются так называемые АЛЬБУМОЗЫ, ПЕПТОНЫ, которые поступают в тонкий кишечник. В тонком кишечнике эти ПОЛИПЕПТИДЫ подвергаются действию целого ряда протеолитических ферментов ПЖЖ: ТРИПСИНОГЕН, ХИМОТРИПСИНОГЕН, ПРОЭЛАСТАЗА (ЭНДОПЕПТИДАЗЫ), ПРОКАРБОКСИПЕПТИДАЗА. Механизм активации всех этих ферментов – ЧАСТИЧНЫЙ ПРОТЕОЛИЗ по каскадному механизму.
ТРИПСИН разрушает внутренние ПЕПТИДНЫЕ связи, в образовании которых принимают участие ЛИЗ и АРГ. ХИМОТРИПСИН разрушает внутренние связи, в образовании которых принимают участие ароматические аминокислоты (ТИР, ТРИ, ФЕН). ЭЛАСТАЗА разрушает внутренние ПЕПТИДНЫЕ связи, в образовании которых принимают участие АЛА, ГЛИ, ПРО, ЦИС. В результате действия ПЕПСИНА, ХИМОТРИПСИНА и ЭЛАСТАЗЫ высокомолекулярные ПЕПТИДЫ расщепляются до низкомолекулярных, которые подвергаются действию ЭКЗОПЕПТИДАЗ, действующих на крайние связи, отщепляя отдельные концевые аминокислоты. КАРБОКСИПЕПТИДАЗА разрушает крайние связи, отщепляя аминокислоты с С-конца. АМИНОПЕПТИДАЗА действует на крайние ПЕПТИДНЫЕ связи со стороны N-конца, отщепляя отдельные аминокислоты. ДИПЕПТИДЫ подвергаются действию ДИПЕПТИДАЗ, продуцирующихся слизистой кишечника сразу в активной форме. Т.о. в результате действия всей этой группы ферментов в ЖКТ белки пищи расщепляются до аминокислот. Образующиеся аминокислоты всасываются стенкой кишечника в кровь и поступают во все органы и ткани. Не всосавшиеся аминокислоты поступают в толстую кишку, где подвергаются процессам гниения. Это процесс бактериального распада веществ под действием микрофлоры или ферментов, вырабатывающихся этой микрофлорой. В норме в толстой кишке подвергаются аминокислоты, а при патологии – белки. Это происходит и в других отделах ЖКТ: в желудке (при понижении кислотности), ротовой полости (при кариесе).
Всасывание – в несколько этапов.
1.За счет вторичного акт. Транспорта с Na+(ионами) конкурируя с углеводами
2.При помощи специфических транслоказ для отдельных групп аминокислот (алифатические,нейтральные, алифатические с развитым скелетом, диаминокарбоновые,серосодержащие)
3.Гаммаглютаминилтранспептидаза раб. в кишечнике и во многих др.тканях, связ. Аминок-ту с карбоксигруппой глутамата, переносит в виде дипептида в клетки, кот. Распадается на восст. Глютатиона (глутамат,цис, глицин).
Аминокислоты в клетке расходуются по порядку значимости: на синтез белков, гормонов,нейромедиаторов,азотсодержащих ген, корнетин, корназин, пурины, пиримидины,холин и т.д.
Нарушение всасывания АК. Различают первичные (при различных формах патологии желудка и кишечника — хронических гастритах, язвенной болезни, раке) и вторичные (функциональные) расстройства секреторной и всасывательной функции эпителия в результате отека слизистой оболочки желудка и кишечника, нарушения переваривания белков и всасывания аминокислот в желудочно-кишечном тракте. Причинами нарушения всасывания аминокислот являются повреждение стенки тонкого кишечника (отек слизистой оболочки, воспаление) или неравномерное по времени всасывание отдельных аминокислот. Это ведет к нарушению (дисбалансу) соотношения аминокислот в крови и синтеза белка в целом, поскольку незаменимые аминокислоты должны поступать в организм в определенных количествах и соотношениях. Чаще всего имеет место нехватка метионина, триптофана, лизина и других аминокислот.
23.Общие пути обмена аминокислот. Реакции трансаминирования, окислительного дезаминирования, непрямого дезаминирования. Роль витамина B6.
1.Трансаминирование (переаминирование) – перенос аминогруппы с α-аминокислоты на α-кетокислоту. Контролируют аминотрансферазы, кофермент фосфоперидоксаль. Проходит в 2 этапа в цитозоли: на первом – фосфоперидоксаль забирает аминогруппу, превращается в перидоксамин, на втором – фосфоперидоксамин передает аминогруппу на α-кетокислоту. В результате образуется новая кетокислота и аминокислота.
2.Окислительное дезаминирование – только для глутамата, так как в клетках активна только глутаматДГ.
- Непрямое дезаминирование АК – состоит из 2-х этапов: переаминирование и окислительное дезаминирование
АЛТ – находится в цитоплазме, наибольшая активность в печени, повышается при различных поражениях ( гепатиты, токс.поражения и т.д.). АСТ – находятся и в цитозоли и в митохондриях, повышается в крови при разрушении клеток более глубоком, характерно для инфаркта. Существует коэффициент Де-Риттиса: отношение АСТ к АЛТ, если больше 1 – то более глуб поражение в митохондриях сердца (инфаркт миокарда), в норме 0,6-0,8; меньше единицы – заболевания печени.
Витамин В6 (пиридоксин, антидерматитный)- самост независ пищевой фактор.Термином вит В6, обознач все три производных 3-оксипиридина, облад-х одинаковой вит активностью: пиридоксин (пиридоксол), пиридоксаль и пиридоксамин:
У члка недостаточность вит В6 встреч редко, хотя некот пеллагроподобные дерматиты, не поддающиеся леч никотиновой к-той,проходят при введении пиридоксина. Все три производных 3-оксипиридина наделены витаминными св-ми, коферментные ф-и выполн только фосфорилированные производные пиридоксаля и пи-ридоксамина.
Участвует в образовании ПФ:
-КОФЕРМЕНТ АМИНОТРАНСФЕРАЗ.
-КОФЕРМЕНТ ДЕКАРБОКСИЛАЗ.
-Принимает участие в ДЕЗАМИНИРОВАНИИ.
-Необходим для образования витамина РР из триптофана.
-Сигма-АМИНОЛЕВУЛИНОВАЯ кислота ® гем.
Т.о. витамин В6 участвует в обмене аминокислот, следовательно, необходим для нормального обмена белков. Суточная потребность-2мг. Источники вит.В6: картофель, пшеница, рис, отруби, печень, дрожжи. Гиповитаминоз приводит к нарушению белкового обмена, что проявляется развитием анемии, дерматита, стоматита, глоссита.
24.Образование и биологические функции биогенных аминов. Обезвреживание биогенных аминов.
Синтез биогенных аминов (нейромедиаторов) из аминокислот связан с вовлечением в метаболизм α-карбоксильной группы аминокислот или ее удалением. Ф-т декарбоксилазы, кофермент ФосфоПеридоксаль (Вит В6). Биогенные амины – часто нейромедиаторы или гормоны.Тирозон->тирамин. Тирозин->диоксифенилаланин->дофамин->норадреналин. Глутамат ->ГАНК (медиатор торможения) улучшает память,ЦНС,выносливость,сост.клетки.
Катепсины – внутриклеточные протеазы находятся в лизосомах, способствуют обновлению белков, убирают дефектные, активируются в кислой среде(оптимум 4,5-5,5), делятся на сириновые,цистеиновые,аспарагиновые.
Реакции ДЕКАРБОКСИЛИРОВАНИЯ – разрушение СООН-группы с выделением СО2. При этом аминокислоты в тканях образуют биогенные амины, которые являются биологически активными веществами (БАВ):
- НЕЙРОМЕДИАТОРОВ (СЕРЕТОНИН, ДОФАМИН, ГАМК),
- Гормоны (АДРЕНАЛИН, НОРАДРЕНАЛИН),
- Регуляторы местного действия ( ГИСТАМИН).
ГАМК является НЕИРОМЕДИАТОРОМ тормозного действия. ДОФАМИН является НЕИРОМЕДИАТОРОМ возбуждающего действия. Он является основой для синтеза АДРЕНАЛИНА и НОР АДРЕНАЛИНА.
ГИСТАМИН повышает секрецию желудочного сока, поэтому применяется в клинической практике при зондировании. Обладает сосудорасширяющим действием, понижает АД.
Дофамин Синтез дофамина происходит в основном в нейронах промежуточного и среднего мозга. Является медиатором дофаминовых рецепторов в подкорковых образованиях ЦНС, в больших дозах расширяет сосуды сердца, стимулирует частоту и силу сердечных сокращений, расширяет сосуды почек, увеличивая диурез.
25.Основные источники аммиака в организме. Непрямое и окислительное дезаминирование аминокислот. Токсичность аммиака. Гипераммониемии.
1.Аммиак постоянно образуется во всех органах и тканях организма. Наиболее активными его продуцентами в кровь являются органы с высоким обменом аминокислот и биогенных аминов – нервная ткань, печень, кишечник, мышцы.
Основными источниками аммиака являются аминокислоты, которые не накапливаются в организме. Избыток высвобождает(теряет) аминогруппу. Следующие реакции: -неокислительное дезаминирование некоторых аминокислот (серина, треонина, гистидина) – в печени, –окислительное дезаминирование глутаминовой кислоты во всех тканях (кроме мышечной), особенно в печени и почках, –дезаминирование амидов глутаминовой и аспарагиновой кислот – в печени и почках, –катаболизм биогенных аминов – во всех тканях, в наибольшей степени в нервной ткани, -жизнедеятельность бактерий толстого кишечника, -распад пуриновых и пиримидиновых оснований – во всех тканях.
- 4. Окислительное. ОКИСЛИТЕЛЬНОМУ ДЕЗАМИНИРОВАНИЮ подвергается только ГЛУ.
ОКИСЛИТЕЛЬНОМУ ДЕЗАМИНИРОВАНИЮ подвергаются и другие аминокислоты, но этот путь является непрямым. Он идёт через ГЛУ и называется процессом НЕПРЯМОГО ОКИСЛИТЕЛЬНОГО ДЕЗАМИНИРОВАНИЯ.
3.Токсичность аммиака
Аммиак является токсичным соединением, находящимся в крови в относительно небольших концентрация, В норме 0,4-0,7 мг/л. Повышение концентрации аммиака (гипераммониемия)до 6 мг – интоксикация, кома, судороги, бред и смерть в тяжелых случаях. При хронической гипераммониемии развивается умственная отсталость.
Аммиак связывается с α-кетоглютаратом, переводит его в глутамат, а далее в глутамин. Это снижает скорость ЦТК, а значит уровень АТФ, что приводит к нарушению работы калий-натриевой АТФ-фазы, не восстанавливается потенциал покоя. Глутамин в норме выходит в кровь, приходит в печень, где отдаёт аммиак на синтез мочевины. Избыток глутамина в мозге приводит к хадержке воды, отеку мозга, в крови гипераммониемия, что снижает щелочные резервы крови ( алкалоз). Снижается ацидоз, возникает алкалоз – это приводит к изм.конформации гемоглобина, что вызывает тканевую гипоксию.
Наследственные и приобретенные формы гипераммониемий
-Приобретенная (вторичные) гипераммониемия развивается вследствие заболеваний печени ивирусных инфекций. В крайне тяжелых случаях она проявляется как тошнота, рвота, судороги, нечленораздельная речь, затуманивание зрения, тремор, нарушение координации движений.
-Наследственные формы гипераммониемии вызваны генетическим дефектом любого из пяти ферментов синтеза мочевины. Соответственно ферменту заболевание делится на пять типов. Первичными признаками гипераммониемий являются сонливость, отказ от пищи, рвота, беспокойство, судороги, нарушение координации движений, тахипноэ, дыхательный алкалоз. Могут развиться печеночная недостаточность, легочные и внутричерепные кровоизлияния.
Наиболее частой является гипераммониемия типа II, связанная с недостатком орнитин-карбамоилтрансферазы. У матери также наблюдается гипераммониемия и отвращение к белковым продуктам. При полном дефекте фермента наследственные имеют раннее начало (в период до 48 часов после рождения).
Основа лечения ограничение белка в диете, позволяет предотвратить нарушения мозговой деятельности.
- Пути обезвреживания аммиака: амидирование, восстановительное аминирование, аммониогенез.
Аммиак – токсичное соединение. Даже небольшое повышение его концентрации оказывает нблагоприятное действие на организм, и прежде всего на ЦНС. Так, повышение концентрации аммиака в мозге до 0,6 ммоль вызывает судороги. К симптомам гипераммониемии относят тремор, нечленораздельную речь, тошноту, рвоту, головокружение, судорожные припадки, потерю сознания. В тяжёлых случаях развивхается кома с летальным исходом
- Синтез мочевины – основной путь обезвреживания аммиака. Связь орнитинового цикла с циклом Кребса. Наследственные нарушения орнитинового цикла.
28 .Взаимосвязь обмена углеводов и липидов. Синтез глюкозы из аминокислот и глицерина. Биосинтез жиров и аминокислот из углеводов. Обмен веществ, или метаболизм, — лежащий в основе жизни закономерный порядок превращения веществ и энергии в живых системах, направленный на их сохранение и самовоспроизведение; совокупность всех химических реакций, протекающих в организме. Все превращения органических веществ, процессы анаболизма и катаболизма тесно связаны друг с другом.На I этапе полисахариды расщепляются до моносахаридов (обычно гексоз); жиры распадаются на глицерин и высшие жирные кислоты, а белки – на составляющие их свободные аминокислоты. Указанные процессы в основном являются гидролитическими, поэтому освобождающаяся в небольшом количестве энергия почти целиком используется организмами в качестве тепла. На II этапе мономерные молекулы (гексозы, глицерин, жирные кислоты и аминокислоты) подвергаются дальнейшему распаду, в процессе которого образуются богатые энергией фосфатные соединения и ацетил-КоА. На III этапе ацетилКоА (и некоторые другие метаболиты, например α-кетоглутарат, оксалоацетат) подвергаются окислению («сгоранию») в цикле ди- и трикарбоновых кислот Кребса. Окисление сопровождается образованием восстановленных форм НАДН + Н+ и ФАДН2.На IV этапе осуществляется перенос электронов от восстановленных нуклеотидов на кислород (через дыхательную цепь). Он сопровождается образованием конечного продукта – молекулы воды. Этот транспорт электронов сопряжен с синтезом АТФ в процессе окислительного фосфорилирования. Для некоторых аминокислот (аланин, аспарагиновая и глутами-новая кислоты) связь с глюконеогенезом является непосредственной, для других она осуществляется через побочные метаболические пути. Следует особо подчеркнуть, что три α-кетокислоты (пируват, оксалоацетат и кетоглутарат), образующиеся соответственно из аланина, аспартата и глутамата, не только служат исходным материалом для синтеза глюкозы, но являются своеобразными кофакторами при распаде ацетильных остатков всех классов пищевых веществ в цикле Кребса для получения энергии. Процесс синтеза углеводов из аминокислот получил название глюконеогенеза. Исходными субстратами для глюконеогенеза являются те аминокислоты, распад которых сопровождается образованием прямо или опосредованно пировиноградной кислоты (например, аланин, серин, треонин и цистеин). Взаимосвязь обмена углеводов, жиров, белков проявляется в двух аспектах: а) в наличии единых промежуточных продуктов обмена и б) во взаимопревращениях углеродов, жиров, белков. Таким образом, процессы распада жиров, белков, углеводов сходятся (в большинстве своем на стадии образования ацетил КоА), образуя в дальнейшем единый метаболический цикл (цикл трикарбоновых кислот), завершающий их превращения. Этим достигается определенная экономия на разнообразии ферментов, на внутриклеточных структурных образованиях, обеспечивающих локализацию ферментных систем и процессов. В условиях истощения углеводных ресурсов организма жиры начинают энергично использоваться в качестве источника энергии. При этом жирные кислоты или непосредственно используются тканями, или превращаются в печени в кетоновые тела, которые поступают в кровь и также утилизируются тканями в качестве энергетического субстрата. Из другого продукта мобилизации жира – глицерина образуется глюкоза , которая поступая в кровь , обеспечивает энергетическим сырьем ткани , предпочитающие глюкозу другим субстратам . При избыточном поступлении в организм углеводов они могут превращаться в жиры . При этом глицерин образуется из промежуточного продукта гликолиза- фосфоглицеринового альдегида , а непосредственным сырьем для синтеза ж.к. является ацетил КоА , образовавшийся при распаде углеводов . Часть аминокислот организма вообще не способен образовывать из других веществ , другие могут быть синтезированы . Но для этого требуется включить в их состав аминогруппу . Источником аминогруппы могут служить другие аминокислоты или свободный аммиак , в свою очередь , образующийся при дизаминировании аминокислот. Т.о. взаимосвязь обмена углеводов, жиров, белков и их взаимопревращения имеет чрезвычайно важное значение для организма человека и животных. Также это обеспечивает возможность создания в организме запаса энергетических субстратов при любом характере питания. Наличие углеводных депо, возможность образования углеводов из продуктов не углеводной природы играет чрезвычайную роль в жизни человека. Благодаря этому обеспечивается относительное постоянство концентрации глюкозы в крови и непрерывное снабжение ею тканей, использующих глюкозу в качестве основного энергетического субстрата (мозговые ткани, почки, эритроциты и пр.).
- Минеральные вещества тканей человека. Регуляция вводно-солевого обмена: антидиуретический гормон, альдостерон и ренин-ангиотензиновая система. Роль почек в регуляции гомеостаза.
Минеральный обмен – совокупность процессов всасывания, усвоения, распределения, превращения и выделения из организма тех веществ, которые находятся в нём преимущественно в виде неорганических соединений. Минеральные вещества в составе биологической жидкости создают внутреннюю среду организма с постоянными физико-химическими свойствами, что обеспечивает нормальное функционирование клеток и тканей. Определения содержания и концентрации ряда минеральных веществ в жидкостях организма является важным диагностическим тестов при многих заболеваниях. В одних случаях нарушение минерального обмена является причиной заболевания, в других – лишь симптомами заболевания, однако любая болезнь в той или иной степени сопровождается нарушением водно-минерального обмена.
По количеству основную часть минеральных соединений организма составляют хлористые, фосфорнокислые и углекислые соли натрия, калия, кальция и магния. Кроме того в организме содержатся соединения железа, марганца, цинка, меди, кобальта, йода и ряда других микроэлементов.
Минеральные соли в водных средах организма частично или полностью растворяются и существуют в виде ионов. Минеральные вещества могут находиться также в форме нерастворимых соединений. В костной и хрящевой тканях сосредоточено 99% всего кальция организма, 87% фосфора, 50% магния. Минеральные вещества входят в состав многих органических соединений, например белков. Минеральный состав некоторых тканей взрослого человека приведён в таблице.
солевой обмен является частью минерального обмена, он представляет собой совокупность процессов поступления в организм воды и солей, главным образом NaCl, распределение их во внутренней среде и выведения из организма. Нормальный водно-солевой обмен обеспечивает постоянный объём крови и других жидкостей организма, осмотическое давление и кислотно-щелочное равновесие. Основным минеральным веществом, благодаря которому организм регулирует осмотическое давление, является натрий, примерно 95% осмотического давления плазмы крови регулируется с помощью этого минерального вещества.
Водно-солевой обмен – это совокупность процессов поступления воды и солей (электролитов) в организм, распределение их во внутренней среде и выведения из организма. Системы регуляции водно-солевого обмена обеспечивают постоянство суммарной концентрации растворённых частиц, ионного состава и кислотно-щелочного равновесия, а также объёма и качественного состава жидкостей организма.
Антидиуретический гормон
Антидиуретический гормон (АДГ), или вазопрессин – пептид с молекулярной массой около 1100 Д, содержащий 9 аминокислот, соединённых одним дисульфидным мостиком.
- Синтез и секреция антидиуретического
гормона
АДГ синтезируется в нейронах гипоталамуса в виде предшественника препрогормона, который поступает в аппарат Гольджи и превращается в прогормон.
Механизм действия
Для АДГ существуют 2 типа рецепторов: V1 и V2. Рецепторы V2, опосредующие главный физиологический эффект гормона, обнаружены на базолатеральной мембране клеток собирательных трубочек и дистальных канальцев – наиболее важных клеток-мишеней для АДГ, которые относительно непроницаемы для молекул воды.
Несахарный диабет
Дефицит АДГ, вызванный дисфункцией задней доли гипофиза, а также нарушениями в системе передачи гормонального сигнала, приводит к развитию несахарного диабета. При этом происходит нерегулируемая экскреция воды, а наиболее опасным последствием является дегидратация организма.
Альдостерон
Альдостерон – наиболее активный минера-локортикостероид, синтезирующийся в коре надпочечников из холестерола.
Синтез и секреция альдостерона клетками клубочковой зоны непосредственно стимулируются низкой концентрацией Na+ и высокой концентрацией К+ в плазме крови. На секрецию альдостерона влияют также простагландины,
Альдостерон не имеет специфических транспортных белков, но за счёт слабых взаимодействий может образовывать комплексы с альбумином. Гормон очень быстро захватывается печенью, где превращается в тетрагидроальдостерон-3-глюкуронид и экскретируется с мочой.
- Механизм действия альдостерона
В клетках-мишенях гормон взаимодействует с рецепторами, которые могут быть локализованы как в ядре, так и в цитозоле клетки. Образовавшийся комплекс гормон-рецептор взаимодействует с определённым участком ДНК и изменяет скорость транскрипции специфических генов. Результат действия альдостерона – индукция синтеза: а) белков-транспортёров Na+ из просвета канальца в эпителиальную клетку почечного канальца; б) Nа+,К+,-АТФ-азы, обеспечивающей удаление ионов натрия из клетки почечного канальца в межклеточное пространство и переносящей ионы калия из межклеточного пространства в клетку почечного канальца; в) белков-транспортёров ионов калия из клеток почечного канальца в первичную мочу; г) митохондриальных ферментов ЦТК, в частности цитратсинтазы, стимулирующих образование молекул АТФ, необходимых для активного транспорта ионов
- В организме взрослого человека содержится в среднем 1000 г кальция. Основным депо кальция в организме (99% всего кальция от общей массы) являются кости. В костях около 99% кальция присутствует в малорастворимой форме кристаллов гидроксиапатита [Са10(РО4)6(ОН)2Н2О]. В виде фосфатных солей в костях находится лишь 1% кальция, который может легко обмениваться и играть роль буфера при изменениях концентрации кальция в плазме крови. Другой фонд кальция (1% от общей массы кальция) – кальций плазмы крови. В плазму крови кальций поступает из кишечника (с водой и пищей) и из костной ткани (в процессе резорбции).
Кальций – не только структурный компонент костной ткани. Ионы кальция играют ключевую роль в мышечном сокращении, увеличивают проницаемость мембраны клеток для ионов калия, влияют на натриевую проводимость клеток, на работу ионных насосов, способствуют секреции гормонов, участвуют в каскадном механизме свёртывания крови.
Паратгормон
Паратгормон (ПТГ) – одноцепочечный полипептид, состоящий из 84 аминокислотных остатков (около 9,5 кД), действие которого направлено на повышение концентрации ионов кальция и снижение концентрации фосфатов в плазме крови.
- Синтез и секреция ПТГ
ПТГ синтезируется в паращитовидных железах в виде предшественника – препрогормона, содержащего 115 аминокислотных остатков.
Секреция ПТГ регулируется уровнем ионов кальция в плазме: гормон секретируется в ответ на снижение концентрации кальция в крови.
- Роль паратгормона в регуляции обмена
кальция и фосфатов
Органы-мишени для ПТГ – кости и почки. В клетках почек и костной ткани локализованы специфические рецепторы, которые взаимодействуют с паратгормоном, в результате чего инициируется каскад событий, приводящий к активации аденилатциклазы.
Кальцитриол
Как и другие стероидные гормоны, кальцитриол синтезируется из холестерола.
ействие гормона направлено на повышение концентрации кальция в плазме крови.
- Строение и синтез кальцитриола
В коже 7-дегидрохолестерол (провитамин D3) превращается в непосредственного предшественника кальцитриола – холекальциферол (витамин D3). В ходе этой неферментативной реакции под влиянием УФ-излучения связь между девятым и десятым атомами углерода в молекуле холестерола разрывается, раскрывается кольцо В, и образуется холекальциферол (рис. 11-38). Так образуется в организме человека большая часть витамина D3, однако небольшое его количество поступает с пищей и всасывается в тонком кишечнике вместе с другими жирорастворимыми витаминами.
- Механизм действия кальцитриола
Кальцитриол оказывает воздействие на тонкий кишечник, почки и кости. Подобно другим стероидным гормонам, кальцитриол связывается с внутриклеточным рецептором клетки-мишени. Образуется комплекс гормон-рецептор, который взаимодействует с хроматином и индуцирует транскрипцию структурных генов, в результате чего синтезируются белки, опосредующие действие кальцитриола. Так, например, в клетках кишечника кальцитриол индуцирует синтез Са2+-переносящих белков, которые обеспечивают всасывание ионов кальция и фосфатов из полости кишечника в эпителиальную клетку кишечника и далее транспорт из клетки в кровь, благодаря чему концентрация ионов кальция во внеклеточной жидкости поддерживается на уровне, необходимом для минерализации органического матрикса костной ткани.
. Рахит
Рахит – заболевание детского возраста, связанное с недостаточной минерализацией костной ткани. Нарушение минерализации кости – следствие дефицита кальция. Рахит может быть обусловлен следующими причинами: недостатком витамина D3 в пищевом рационе, нарушением всасывания витамина D3 в тонком кишечнике, снижением синтеза предшественников кальцитриГола из-за недостаточного времени пребывания на солнце, дефектом 1α-гидроксилазы, дефектом рецепторов кальцитриола в клетках-мишенях.
РОЛЬ КАЛЬЦИТОНИНА В РЕГУЛЯЦИИ ОБМЕНА КАЛЬЦИЯ
Кальцитонин – полипептид, состоящий из 32 аминокислотных остатков с одной дисульфидной связью. Гормон секретируется парафолликулярными К-клетками щитовидной железы или С-клетками паращитовидных желёз в виде высокомолекулярного белка-предшественника. Секреция кальцитонина возрастает при увеличении концентрации Са2+ и уменьшается при понижении концентрации Са2+ в крови. Кальцитонин – антагонист паратгормона. Он ингибирует высвобождение Са2+ из кости, снижая активность остеокластов. Кроме того, кальцитонин подавляет канальцевую реабсорбцию ионов кальция в почках, тем самым стимулируя их экскрецию почками с мочой. Скорость секреции кальцитонина у женщин сильно зависит от уровня эстрогенов. При недостатке эстрогенов секреция кальцитонина снижается. Это вызывает ускорение мобилизации кальция из костной ткани, что приводит к развитию остеопороза.
31)Нейро-гуморальная регуляция обмена веществ. Основные механизмы регуляции обмена веществ на клеточном уровне. Роль гормонов в регуляции обмена веществ.
Гормо ны-биологически активные сигнальные химические вещества, выделяемые эндокринными железами непосредственно в организме и оказывающие дистанционное сложное и многогранное воздействие на организм в целом либо на определённые органы и ткани-мишени. Гормоны служат гуморальными (переносимыми с кровью) регуляторами определённых процессов в различных органах и системах. Используются в организме для поддержания его гомеостаза, а также для регуляции многих функций (роста, развития, обмена веществ, реакции на изменения условий среды).
Гормонов имеющих белковую или полипептидную природу называют тропинами, так как они оказывают направленное стимулирующее действие на процессы роста и обмена веществ организма и на функцию периферических эндокринных желез. Обмен углеводов, липидов и аминокислот главным образом регулируют инсулин, глюкагон, кортизол и адреналин.Универсальным энергетическим субстратом для всех тканей является глюкоза, причем ее уровень в крови контролирует синтез и секрецию гормонов поджелудочной железы – инсулина и глюкагона. Метаболизм углеводов, жиров и белков абсорбтивный период.При пищеварении в крови повышаются концентрации глюкозы, аминокислот, хиломикронов и ЛПОНП. Увеличение уровня глюкозы в крови стимулирует секрецию гормона инсулина β-клетками поджелудочной железы. Метаболизм углеводов, жиров и белков постабсорбтивный период и при голодании.
1 фаза голодания. В течение постабсорбтивного периода и при дальнейшем голодании уровень глюкозы в крови постепенно снижается, поэтому концентрация инсулина уменьшается и повышается концентрация глюкагона. Индекс инсулин/глюкагон снижается. 2 фаза голодания характеризуется дальнейшим снижением индекса инсулин/глюкагон, вызванным ускорением синтеза и секреции глюкагона. Кроме того, продолжает повышаться синтез и секреция другого гормона – кортизола. Глюкагон стимулирует липолиз в жировой ткани. Это приводит к повышению концентрации жирных кислот. Глюкоза не поступает в мышцы из-за низкой концентрации инсулина, поэтому эта ткань получает энергию за счет окисления жирных кислоты и кетоновых тел. В крови повышается концентрация мочевины и увеличивается ее содержание в моче. Это является следствием ускорения катаболизма мышечных белков и дезаминирования аминокислот в печени под влиянием кортизола. 3 фаза наступает при продолжении голодания более недели. В этом случае основными энергетическими субстратами становятся жирные кислоты и кетоновые тела. Скорость катаболизма белков и глюконеогенез из аминокислот тормозится, вследствие чего синтез и выведение мочевины из организма уменьшаются. Снижение количества белков сопровождается атрофией тканей.
32)Классификация гормонов. Иерархия регуляторных систем. Нейропептиды мозга: либерины и статины. Тропные гормоны гипофиза. Эндокринные заболевания. Гормоны можно классифицировать, исходя из разных критериев: По растворимости (гидрофильные и липофильные). По химической структуре: 1.производные аминокислот: производные тирозина: тироксин, трийодтиронин, дофамин, адреналин, норадреналин; 2. белково-пептидные гормоны: полипептиды: глюкагон, кортикотропин, меланотропин, вазопрессин, окситоцин, пептидные гормоны желудка и кишечника; простые белки (протеины): инсулин, соматотропин, пролактин, паратгормон, кальцитонин; сложные белки (гликопротеиды): тиреотропин, фоллитропин, лютропин. стероидные гормоны: кортикостероиды (альдостерон, кортизол, кортикостерон); половые гормоны: андрогены (тестостерон), эстрогены и прогестерон. 3. По механизму передачи сигнала или по расположению рецепторов. По этому критерию гормоны делятся на 2 основные группы. К первой группе относятся стероиды, иодтиронины и кальцитриол. Рецепторы этих гормонов располагаются в цитоплазме или на ядре. Ко второй группе относятся водорастворимые гормоны, которые взаимодействуют с рецепторами, находящимися на плазматической мембране. 4. По природе сигнала, опосредующего гормональный внутриклеточный эффект. 5.по биологическим функциям — гормоны, регулирующие обмен углеводов, липидов и аминокислот (инсулин, глюкагон, кортизол, адреналин), регулирующие водно-солевой об прессин, альдостерон), обмен кальция (паратгормон, кальцитриол, кальцитонин), регулирующие репродуктивную функцию (эстрадиол, тестостерон, прогестерон). Тропные гормоны -либерины и статины гипоталамуса, некоторые гормоны гипофиза) регулируют синтез и других гормонов.
Статины и либерины называются рилизинг-гормонами. От их концентрации зависит деятельность гипофиза, а значит и функция периферических эндокринных желез (надпочечников, щитовидной, яичников или яичек).В настоящее время идентифицированы следующие статины и либерины:гонадолиберины (фоллиберин и люлиберин) ;соматолиберин;пролактолиберин;тиролиберин; меланолиберин; кортиколиберин; соматостатин; пролактостатин (дофамин);меланостатин.
Тропные гормоны (син. тропины) – это гормоны, вырабатываемые передней долей гипофиза, регулирующие деятельность остальных желез внутренней секреции (щитовидной железы, надпочечников, яичников и яичек) и опосредованно влияющие на многие процессы жизнедеятельности организма.
К ним относятся адренокортикотропный гормон, контролирующий функционирование коры надпочечников;тиреотропный гормон, регулирующидеятельность щитовидной железы;соматотропный гормон (или гормон роста), регулирующий рост и развитие организма;липотропные гормоны, ускоряющие расщепление жиров и тормозящие отложения жира;меланоцитстимулирующий гормон, отвечающий за образование и отложение в волосах и коже пигмента меланина;пролактинлактогенный гормон), отвечающий за выработку молока в период лактации;а также группа гонадотропных гормонов: фолликулостимулирующий гормон (ФСГ), лютеинизирующий гормон (ЛГ), отвечающдеятельность половых желез.
Секреция и выброс тропных гормонов гипофиза осуществляется под контролем гипоталамуса и вырабатываемых им гипоталамических нейрогормонов (веществ, стимулирующих или тормозящих процессы секреции в гипофизе).
Избыточная выработка тропных гормонов может быть обусловлена аденомой гипофиза или его гиперплазией и приводит к развитию таких эндокринных заболеваний, как акромегалия, болезнь Иценко – Кушинга. Недостаточная же выработка становится причиной развития гипоталамо-гипофизарной недостаточности.
33)Гормоны щитовидной железы,влияние на обмен в-в.Гипо и гипертиреозы
Щитовидная железа состоит из двух долей, соединенных перешейком и расположенных на шее по обеим сторонам трахеи ниже щитовидного хряща. Она имеет дольчатое строение. Ткань железы состоит из фолликулов, заполненных коллоидом, в котором имеются йодсодержащие гормоны тироксин (тетрайодтиронин) и трийодтиронин в связанном состоянии с белком тиреоглобулином. В межфолликулярном пространстве расположены парафолликулярные клетки, которые вырабатывают гормон тиреокальцитонин. Содержание тироксина в крови больше, чем трийодтиронина. Однако активность трийодтиронина выше, чем тироксина. Эти гормоны образуются из аминокислоты тирозина путем ее йодирования. Инактивация происходит в печени посредством образования парных соединений с глюкуроновой кислотой. Кальцитонин, или тиреокальцитонин, вместе с паратгормоном околощитовидных желез участвует в регуляции кальциевого обмена. Под его влиянием снижается уровень кальция в крови (гипокальциемия). Это происходит в результате действия гормона на костную ткань, где он активирует функцию остеобластов и усиливает процессы минерализации. Функция остеокластов, разрушающих костную ткань, напротив, угнетается. В почках и кишечнике кальцитонин угнетает реабсорбцию кальция и усиливает обратное всасывание фосфатов. Продукция тиреокальцитонина регулируется уровнем кальция в плазме крови по типу обратной связи. При снижении содержания кальция тормозится выработка тиреокальцитонина, и наоборот.
Йодсодержащие гормоны выполняют в организме следующие функции: 1) усиление всех видов обмена (белкового, липидного, углеводного), повышение основного обмена и усиление энергообразования в организме; 2) влияние на процессы роста, физическое и умственное развитие; 3) увеличение частоты сердечных сокращений; 4) стимуляция деятельности пищеварительного тракта: повышение аппетита, усиление перистальтики кишечника, увеличение секреции пищеварительных соков; 5) повышение температуры тела за счет усиления теплопродукции; 6) повышение возбудимости симпатической нервной системы.
Нарушения функции щитовидной железы проявляются ее гипофункцией и гиперфункцией. Если недостаточность функции развивается в детском возрасте, то это приводит к задержке роста, нарушению пропорций тела, полового и умственного развития. Такое патологическое состояние называется кретинизмом. У взрослых гипофункция щитовидной железы приводит к развитию патологического состояния – микседемы. При этом заболевании наблюдается торможение нервно-психической активности, что проявляется в вялости, сонливости, апатии, снижении интеллекта, нарушении половых функций, угнетении всех видов обмена веществ и снижении основного обмена. У таких больных увеличена масса тела за счет повышения количества тканевой жидкости и отмечается одутловатость лица. щитовидной железы может развиться у людей проживающих в местностях, где в воде и почве отмечается недостаток йода. Это так называемый эндемический зоб. При гиперфункции щитовидной железы развивается заболевание тиреотоксикоз (диффузный токсический зоб, Базедова болезнь, болезнь Грейвса). Характерными признаками этого заболевания являются увеличение щитовидной железы (зоб) экзофтальм, тахикардия, повышение обмена веществ, особенно основного, потеря массы тела, увеличение аппетита, нарушение теплового баланса организма, повышение возбудимости и раздражительности. Недостаточность поступления йода приводит к развитию гипотиреоза — снижения продукции гормонов щитовидной железы. Данное состояние может развиться как до, так и во время вынашивания ребенка, приводя к осложнениям. В течение беременности возможны такие проблемы, как угроза прерывания, самопроизвольные выкидыши, анемия, гестоз, мертворождения
34)Инсулин и глюкагон,хим природа и механизм дей-ия на углеводный и липидный обмен.Сахарный диабет
Поджелудочная железа относится к железам со смешанной функцией. Эндокринная функция осуществляется за счет продукции гормонов панкреатическими островками (островками Лангерганса). В островках имеется несколько типов клеток.
a -клетки, вырабатывающие инсулинb-клетки продуцируют инсулин, d-клетки синтезируют соматостатин, который угнетает секрецию инсулина и глюкагона. G-клетки вырабатывают гастрин. Основную массу составляют b -клетки вырабатывают глюкагон,
- Инсулин влияет на все виды обмена веществ, но, прежде всего, на углеводный. Под воздействием инсулина происходит уменьшение концентрации глюкозы в плазме крови (гипогликемия). Это связано с тем, что инсулин способствует превращению глюкозы в гликоген в печени и мышцах (гликогенез). Он активирует ферменты, участвующие в превращении глюкозы в гликоген печени, и ингибирует ферменты, расщепляющие гликоген. Инсулин также повышает проницаемость клеточной мембраны для глюкозы, что усиливает ее утилизацию. Инсулин стимулирует синтез белка из аминокислот и уменьшает катаболизм белка. Инсулин регулирует жировой обмен. Образование инсулина регулируется уровнем глюкозы в плазме крови. Гипергликемия способствует увеличению выработки инсулина, гипогликемия уменьшает образование и поступление гормона в кровь. Некоторые гормоны желудочно-кишечного тракта, такие как желудочный ингибирующий пептид, холецистокинин, секретин, увеличивают выход инсулина. Антагонистами инсулина по характеру действия на углеводный обмен являются глюкагон, АКТГ, соматотропин, глюкокортикоиды, адреналин, тироксин. Введение этих гормонов вызывает гипергликемию. Недостаточная секреция инсулина приводит к заболеванию, которое получило название сахарного диабета.Симптомы частое мочеиспускание и чувство неутолимой жажды, приводящие к обезвоживанию организма;быстрая потеря веса, зачастую несмотря на постоянное чувство голода;ощущение слабости или усталости;неясность зрения (“белая пелена” перед глазами);сложности с половой активностью;онемение и покалывание в онемевших конечностях ощущение тяжести в ногах;головокружения;медленное излечивание инфекционных заболеваний;медленное заживление ран;падение температуры тела ниже средней отметки;быстрая утомляемость;судороги икроножных мышц;кожный зуд и зуд в промежности;фурункулез;боль в области сердца.
.a клетки островков Лангерганса синтезируют глюкагон, который является антагонистом инсулина. Под влиянием глюкагона происходит распад гликогена в печени до глюкозы. В результате этого повышается содержание глюкозы в крови. Глюкагон способствует мобилизации жира из жировых депо. Секреция глюкагона также зависит от концентрации глюкозы в крови. Гипергликемия тормозит образование глюкагона, гипогликемия, напротив, увеличивает
35)Метаболические ф-ии печени.Роль печени в обмене углеводов,липидов и белков.Липотропные факторы. . Реакция обезвреживания веществ в печени: окисление и коньюгация. Метаболизм чужеродных и лекарственных соединений. Химический канцерогенез.
Метаболизм углеводов
Конечными продуктами расщепления углеводов, поступающих в организм человека, являются глюкоза, фруктоза и галактоза, В печени фруктоза и галактоза превращаются в глюкозу, поэтому метаболизм глюкозы является общим конечным путем метаболизма всех углеводов.
Во всех клетках энергия запасается в виде АТФ, который образуется в ходе анаэробного (гликолиз) или аэробного (цикл лимонной кислоты) расщепления глюкозы. В печени и жировой ткани расщепление глюкозы может также происходить по пентозофосфатному пути, что позволяет не только получить энергию, но и выработать кофактор, играющий важную роль в синтезе жирных кислот. Глюкоза, поступающая в кровь после еды, запасается в организме в форме гликогена. Если депо гликогена насыщены, то поступающая глюкоза превращается в жир. Гликоген является легкодоступным источником глюкозы.
Печень обладает уникальной способностью вырабатывать большое количество глюкозы из молочной и пировиноградной кислоты, аминокислот (в основном из аланина) и глицерола (образуемого в ходе метаболизма жиров). Нормальная концентрация глюкозы в крови поддерживается за счет глюконеогенеза в печени. Глюкокортикоиды, кате-холамины, глюкагон и тиреоидные гормоны потенциируют глюконеогенез, в то время как инсулин, напротив, ингибирует.
Метаболизм жиров
При насыщении депо углеводов избыток поступающих с пищей жиров (и белков) превращается в печени в жиры. Образующиеся жирные кислоты могут немедленно использоваться в качестве источника энергии или откладываться в жировой ткани или печени. В качестве источника энергии почти все клетки организма непосредственно утилизируют жирные кислоты, образующиеся из жиров пищи или синтезируемые в ходе промежуточного метаболизма углеводов и белков. Исключением являются эритроциты и мозговое вещество почки, где может утилизироваться только глюкоза. Нейроны в качестве источника энергии в обычных условиях используют только глюкозу, но через несколько дней голодания могут переключаться на жирные кислоты.Жирные кислоты, образующиеся из жиров, вначале окисляются до ацетилкоэнзима А (ацетил-KoA), который, в свою очередь, окисляется в цикле лимонной кислоты с образованием АТФ. Печень обладает высокой способностью к окислению жирных кислот, в результате чего из избытка ацетил-KoA образуется ацетоацетат. Ацетоацетат, высвобождаемый гепатоцитами, служит альтернативным и легкодоступным (ацетоацетат быстро превращается в ацетил-КоА) циркулирующим в крови энергетическим субстратом для других видов клеток. Глюкагон усиливает окисление жирных кислот, а инсулин ингибирует его.Ацетил-КоА, кроме того, используется печенью для образования холестерина и фосфолипидов, необходимых для синтеза клеточных мембран во всем организме. Синтезируемые в печени липопротеины имеют важное значение для транспорта липидов в крови.
Метаболизм белков
Печень играет ключевую роль в метаболизме белков. Если печень прекращает участвовать в метаболизме белков, то через несколько дней наступает смерть. Метаболизм белков осуществляется в несколько последовательных этапов: 1) дезаминиро-вание аминокислот; 2) образование мочевины (для элиминации аммиака, образующегося при дезами-нировании аминокислот); 3) взаимопревращения между заменимыми аминокислотами; 4) синтез белков плазмы. Дезаминирование необходимо для превращения избытка аминокислот в углеводы и жиры. В ходе ферментативных процессов (чаще всего трансами-нирование) аминокислоты превращаются в соответствующие кетокислоты, а в качестве побочного продукта реакции образуется аммиак. Дезаминирование аланина весьма важно для глюконеогенеза в печени.
Печень играет важную роль в метаболизме гормонов, витаминов и минеральных веществ. Функция щитовидной железы зависит от образования в печени из тироксина (T4) более активного трийод-тиронина (T3). Метаболизм тиреоидных гормонов происходит преимущественно в печени. Печень является основным местом метаболической деградации инсулина, стероидных гормонов (эстроген, альдостерон, кортизол), глюкагона и антидиуретического гормона. Гепатоциты являются основным местом накопления витаминов A, B12, E и D. Наконец, в печени образуется трансферрин и гаптогло-бин, играющие важную роль в метаболизме железа, а также церулоплазмин, необходимый для метаболизма меди.
- Биохимия крови. Основные биохимические константы крови. Белки плазмы крови. Ферменты крови. Липопротеины крови. Биосинтез гема. Особенности обмена в эритроцитах.
37) Биохимия соединительной ткани. Коллаген, эластин – особенности строения и химического состава. Роль витамина С в обмене коллагена. Наследственные нарушения обмена соединительной ткани.
Соединительная ткань составляет примерно 50% от массы тела. Рыхлая соединительная ткань подкожной клетчатки, компактная кость и зубы, сухожилия и межмышечные фасциальные прослойки, кожа и внутриорган-ная строма паренхиматозных органов, нейроглия и брюшина – все это соединительная ткань.Эластин – основной белковый компонент, из которого состоят эластические волокна. Он отличается от коллагена по химическому составу и молекулярной структуре. Общими для эластина и коллагена являются большое содержание глицина и пролина, наличие оксипролина, хотя последнего в эластине примерно в 10 раз меньше, чем в коллагене. Как и в коллагене, в эластине мало метионина и отсутствуют триптофан и цистеин. Наиболее изучена роль витамина С в синтезе коллагена . При авитаминозе С (цинге) нарушается гидроксилирование проколлагена и образование тройной спирали коллагена. Кроме того, происходит угнетение синтеза коллагена и протеогликанов, возможно, обусловленное инактивацией ИФР-I .
38) Особенности химического состава и обмена веществ в костной ткани. Белки костной ткани. Рахит и витамин D.
Химический состав кости зависит от состояния исследуемой кости, возрастных и индивидуальных особенностей. Свежая кость взрослого человека содержит 50 % воды; 15,75 % жира; 12,25 % органических веществ и 22 % неорганических веществ. Высушенная и обезвоженная кость содержит примерно 2/3 неорганического вещества и 1/3 — органического. Если обработать кость кислотой, т. е. произвести декальцинацию, то минеральные соли удаляются. Такая кость, состоящая только из одного органического вещества, сохраняет все детали формы, но отличается чрезвычайной гибкостью и эластичностью. Так, у детей кости гораздо беднее минеральными веществами (неорганическими), поэтому отличаются большей гибкостью и меньшей твердостью. Рахит может возникать при хронических заболеваниях почек, при которых развивается хроническая почечная недостаточность. Также может наблюдаться при хронических заболеваниях печени и обструкции желчевыводящих путей, когда нарушается образование или поступление желчи в кишечник и усвоение жирорастворимых витаминов, в частности витамина Д. Рахит сопровождается вялостью, мышечной слабостью, задержкой роста, деформации трубчатых костей и зубов. Недостаток витамина Д приводит к уменьшению кальция и всасывания фосфора, что является первопричиной деминерализации кости.
39) Ткани зуба. Особенности состава и метаболизма.
Эмаль имеет следующий состав: неорганические вещества – 95%, органические – 1.2%, вода – 3,8%. Неорганические вещества: хлорапатит – 4.39%, фторапатит – 0,63% , гидроксиапатит – 75,04%, карбонапатит -12,06%, карбонат кальция – 1,33%, карбонат магния -1,62%. В составе химических неорганических соединений кальций составляет 37%, а фосфор – 17%. Состояние эмали зуба во многом определяется соотношением Са/Р как элементов, составляющих основу эмали зуба. Здоровая эмаль молодых людей имеет более низкий коэффициент Са/Р, чем эмаль зубов взрослых, этот показатель уменьшается также при деминерализации эмали. Для апатитов, каковыми являются кристаллы эмали зубы, молярное соотношение Са/Р составляет 1,67. Соотношение этих компонентов может изменяться как в сторону уменьшения (1,.33), так и в сторону увеличения (2,0). Соотношение Са/Р в кристаллах в норме вальирует от 1,3 до 2,0. При повышении этого коэффициента устойчивость эмали возрастает.
40) Особенности состава и метаболизма гингивальной жидкости и слюны.
Ротовая жидкость – это биологическая жидкость, которая кроме секрет слюнных желез, включает микрофлору и продукты ее жизнедеятельности, содержимое пародонтальных карманов, десневую жидкость, десквамированный эпителий, распад мигрирующих в полость рта лейкоцитов, остатки пищевых продуктов и т.д. Десневая жидкость, постоянно поступая в полость рта из десневой бороздки или пародонтального кармана, противодействует смещению реакции среды в зубном налете, камне и ротовой жидкости. рН десневой жидкости колеблется в среднем от 7,9 до 8,3. Такие значения поддерживаются высоким уровнем мочевины и аммиака. Слюна является наименее изученной и самой недооцененной из всех жидкостей организма. Тем не менее, этот небольшой по объему секрет играет жизненно важную роль в сохранении интеграции тканей полости рта. У взрослого человека за сутки выделяется 1,5—2 л слюны всеми слюнными железами, если учитывать, что вес их равен 65 г, станет очевидным представление об интенсивности обмена веществ.